靳英华 , 许嘉巍, 宗盛伟, 王鹏
, 许嘉巍, 宗盛伟, 王鹏
东北师范大学地理科学学院,吉林 长春 130024
JIN Ying-hua, XU Jia-wei, ZONG Sheng-wei, WANG Peng
中图分类号: Q948
文献标识码: A
文章编号: 1000-0690(2014)12-1526-07
通讯作者:
收稿日期: 2013-09-20
修回日期: 2014-03-20
网络出版日期: 2014-12-10
版权声明: 2014 地理科学 《地理科学》杂志 版权所有
基金资助:
作者简介:
作者简介:靳英华(1968-),女,吉林长春人,博士,副教授,主要从事自然地理学研究。E-mail: jinyh796@nenu.edu.cn
展开
摘要
为了探讨氮沉降增加对苔原植被的影响,特别是对草本植物侵入苔原的作用,在长白山高山苔原带进行了连续4 a的人工氮沉降模拟实验,测定3种设定的氮沉降水平下牛皮杜鹃(Rhododendron chrysanthum)、笃斯越橘(Vaccinium uliginosum)和小叶章(Deyeuxia angustifolia)的生长状况和群落结构变化。研究结果表明:① 与牛皮杜鹃、笃斯越橘等苔原本地种相比,氮沉降量增加更有利于侵入的小叶章生长;牛皮杜鹃生长对氮沉降量变化响应微弱,氮沉降量增加能明显抑制笃斯越橘的生长。② 无小叶章侵入的牛皮杜鹃和笃斯越橘斑块在不同氮沉降量条件下,群落结构变化不明显,氮沉降增加不是小叶章侵入苔原带的直接原因。③ 在小叶章侵入牛皮杜鹃和笃斯越橘斑块后,氮沉降量增加强化了小叶章的竞争能力,逐渐取代牛皮杜鹃或笃斯越橘,成为优势种,推动高山苔原向高山草甸转化。因此,随着氮沉降量的不断增加,长白山苔原带将面临退化与草甸化。
关键词:
Abstract
In order to explore the effects of increased nitrogen deposition on alpine tundra vegetation change, especially the herb invasion, in Changbai Mountains, the experiments of simulated increased nitrogen deposition were conducted over four years in the alpine tundra zone of the Changbai Mountains. The experiment included three nitrogen deposition treatments. The changes of growth and community structure were measured for Rhododendron chrysanthum, Vaccinium uliginosum and Deyeuxia angustifolia. The main results were as follows: 1) Compared to the Rhododendron chrysanthum, Vaccinium uliginosum and other tundra native species, the increase of nitrogen deposition is more conducive to the growth of Deyeuxia angustifolia, it was no significant response to increase of nitrogen deposition for growth of Rhododendron chrysanthum, it was significantly suppressed by increase of nitrogen deposition for growth of Vaccinium uliginosum. 2) The structure of Rhododendron chrysanthum and Vaccinium uliginosum community with no Deyeuxia angustifolia invasion did not change obviously. That shows that the increase of nitrogen deposition is not the direct reason of invasion of Deyeuxia angustifolia to tundra communities. 3) When Deyeuxia angustifolia invades the Rhododendron chrysanthum and Vaccinium uliginosum community, the increased nitrogen deposition contributes to the competitive ability of Deyeuxia angustifolia. And it promotes the transformation of alpine tundra to alpine meadow. With the increasing amount of nitrogen deposition, the tundra zone of the Changbai Mountains may face degradation and transform into tundra meadow.
Keywords:
以大气氮素沉降增加为主要特征的全球变化正在改变着人类赖以生存的生态环境,严重威胁着陆地生态系统的健康[1~6]。在工业化前,NOy和NHx沉降主要发生在热带地区,而目前北半球温带地区的NOy和NHx沉降量较工业化前增加4倍多,沉降数量大大超过了热带地区[7~10]。中国一些地区已存在着高氮沉降问题,大部分湿润地区氮沉降量(N)已经超过20 kg/ (hm2·a)。在南方森林区,氮沉降量已高达30~73 kg/ (hm2·a),与欧洲最高的氮沉降量相当[11~16]。氮沉降量在未来几十年将继续增加,还将呈现出全球化的趋势,预计到2050年氮沉降量即可翻番,随着中国经济的进一步发展,氮沉降问题也将越来越严重[17,18]。
各种植物对氮输入增加的响应并不一致,许多优势种的生长不受氮浓度的影响,而某些亚优势种,特别是草本植物,其生长量与丰富度会随有效氮数量的增加而增加,由此改变了植物群落的结构[19~21]。欧洲石楠(Heather)林之所以逐渐转变成草原,是氮沉降增加导致草本植物( 如羽状短柄草Brachy podium pinnatum)竞争能力增强的结果[22]。美国加州南部一年生草本植物正逐步取代海滨山艾树灌木,多数观点认为是放牧和火烧造成的,但最近越来越多的学者认为大气氮沉降可能起了重要作用[22]。Bobbink通过实验,证实了氮沉降对长有小灌木的草地植被组成的影响[23] 。
因贫氮生境中部分物种随着氮沉降的增加生长受到抑制[24],高山苔原作为贫氮区生态系统对氮沉降的增加更为敏感[25],Bowman 等通过对Niwot Ridge高山苔原的观察证实,在人类活动很少的高山苔原区氮沉降也是增加的,由于氨态和硝态氮沉降的混合影响,苔原的生物量出现增加,然而氮沉降的增加对植被组成、物种丧失的影响还不清楚[26]。 对瑞典北方亚高山苔原进行的施肥实验表明,苔原优势种生长缓慢的小灌木岩高兰(Empetrum hermaphroditum)被快速生长的草本曲芒发草(Deschampsia flexuosa)代替[27]。
长白山高山苔原是中国湿润区唯一的苔原,植被以矮小灌木如牛皮杜鹃(Rhododendron chrysanthum)、笃斯越橘(Vaccinium uliginosum)为优势物种。然而氮沉降增加对长白山苔原植被的影响研究目前未见报告。鉴于近年来苔原带出现了严重的小叶章(Deyeuxia angustifolia)等草本植物斑块状侵入现象[28,29],本研究试图通过人工施氮模拟不同氮沉降水平下的牛皮杜鹃、笃斯越橘和小叶章的生长状况,观察群落结构的变化,探讨氮沉降对长白山高山苔原植被变化特别是草本植物侵入的作用。
实验区选在长白山西坡苔原带(41°59.515′N 、128°.913′E),海拔2 218~2 222 m,面积为100 m×50 m。实验区地表多为碱性粗面岩风化物和少量火山灰,地貌为流水改造的火山锥坡面,土壤为薄层山地苔原土,气候为寒带气候。实验区植物中极地或高山种属约占80%,以灌木为主,牛皮杜鹃群落和笃斯越橘群落为苔原带最常见的植物群落,群落中存在2个层片,为灌木层片、苔藓-地衣层片,小叶章等草本植物较少,未形成草本植物层片。
小叶章为禾本科野青茅属的多年生、湿中生根茎禾草,多分布于疏林和低湿地,在长白山西坡的岳桦林(Betula emanii)下尤为常见。长白山西坡的小叶章侵入苔原带始于20世纪80年代末,首先出现在苔原带下缘(林线附近),并逐年增多、上侵,形成小叶章占优势的斑块[28]。
本研究于2010~2013年进行,以4 a为研究周期。实验分为二组,分别为牛皮杜鹃组和笃斯越橘组。牛皮杜鹃组包括:无小叶章侵入斑块(小叶章多度0)、小叶章轻度侵入斑块(小叶章多度0.20~0.30)、小叶章中度侵入斑块(小叶章多度0.45~0.55)、小叶章重度侵入斑块(小叶章多度0.70~0.80)和完全侵入斑块(小叶章多度0.95以上);笃斯越橘组包括:无小叶章侵入斑块(小叶章多度0)、小叶章轻度侵入斑块(小叶章多度0.20~0.30)、小叶章中度侵入斑块(小叶章多度0.45~0.55)、小叶章重度侵入斑块(小叶章多度0.70~0.80)。试验设计参照欧洲NITREX项目和北美Harvard Forest等类似研究的设计[30],每组试验氮沉降为3个处理:高、中沉降量和对照(自然沉降)。在每年6~8月的月初,以喷淋方式施用硝酸铵,共3次,每次施用硝酸铵为14.3 g/m2 和7.2 g/m2及0 g/m2,每年施纯氮为15 g/m2、7 g/m2和0 g/m2;每种处理均设4个样方重复,样方面积为2 m×2 m,共计108个样方。
每年9月初进行样方调查,计数样方中1 m×1 m的面积内的物种数量、株数、株高,利用ADC多光谱冠层指数测量仪测量盖度和NDVI指数,利用LAI-2200冠层仪测叶面积指数。2013年9月初全面测量,除历年测量的指标外,又测定了代表平均生长状况样方内的地上、地下生物量。
由于牛皮杜鹃、笃斯越橘和小叶章的芽位不同,直接对比生长季的高度变化和生物量变化无意义。本研究采用相对高度和相对生物量,可以反映随着氮沉降量的增加,植株高度和生长量的差异。其中,相对生长高度=HX/H1;相对生长量=GX/G1(X表示中、高沉降量的高度或生物量,1表示自然沉降量的高度或生物量)。
根据群落结构在4 a实验中的变化程度,将斑块分为2种类型。其中,无小叶章侵入的牛皮杜鹃斑块(简称牛皮杜鹃斑块)、无小叶章侵入的笃斯越橘斑块(简称笃斯越橘斑块)和完全侵入的小叶章斑块(简称小叶章斑块)的群落结构变化不明显,称为稳定斑块;其余斑块的群落结构变化明显,称为不稳定斑块,不稳定斑块包括牛皮杜鹃组和笃斯越橘组的小叶章轻度侵入斑块、中度侵入斑块和重度侵入斑块。
通过4 a连续的模拟氮沉降观测,随着氮沉降量的增加,牛皮杜鹃、笃斯越橘和小叶章3种稳定斑块的物种组成都没发生变化,但与施肥前相比3种稳定斑块的植株数量和盖度的变化存在一定差异。其中,牛皮杜鹃斑块的3个氮沉降处理中牛皮杜鹃植株数量和盖度差异小(P>0.05),笃斯越橘斑块的高氮处理中笃斯越橘植株数量和盖度与无氮添加的对照相比显著减少 (P <0.05),而小叶章斑块随着氮沉降量的增加小叶章植株数量和盖度显著增加 (P< 0.05)(表1)。
3种稳定斑块中牛皮杜鹃斑块的NDVI指数和叶面积指数最大,小叶章斑块的NDVI指数和叶面积指数最小。中、高氮沉降量处理的牛皮杜鹃斑块NDVI指数和叶面积指数与自然沉降量的对照相比差异不显著,中沉降量的NDVI指数和叶面积指数略高,高沉降量的NDVI指数和叶面积指数略低 (P>0.05)。随着氮沉降量的增高,笃斯越橘斑块的NDVI指数和叶面积指数明显降低(P<0.05),而小叶章斑块相反,其NDVI指数和叶面积指数显著增高(P<0.05) (表1)。
表1 不同氮沉降量的稳定斑块变化
Table 1 Changes in the three stable patches with different nitrogen deposition
| 处理 | 牛皮杜鹃斑块 | 笃斯越橘斑块 | 小叶章斑块 | |
|---|---|---|---|---|
| 植株数量(株) | 自然沉降量 | 18±0.57a | 36±0.84a | 84±0.78a |
| 中氮沉降量 | 18±0.69a | 35±0.87a | 103±0.89b | |
| 高氮沉降量 | 17±0.63a | 28±0.53b | 120±0.94c | |
| 盖度(%) | 自然沉降量 | 84±0.45a | 82±0.48a | 80±0.28a |
| 中氮沉降量 | 85±0.38a | 82±0.36a | 86±0.32b | |
| 高氮沉降量 | 86±0.52a | 78±0.49b | 93±0.43c | |
| NDVI指数 | 自然沉降量 | 0.832±0.008a | 0.785±0.004a | 0.564±0.003a |
| 中氮沉降量 | 0.844±0.004a | 0.749±0.005b | 0.624±0.004b | |
| 高氮沉降量 | 0.821±0.005a | 0.695±0.006c | 0.682±0.001c | |
| 叶面积指数 | 自然沉降量 | 4.51±0.031a | 3.88±0.019a | 3.55±0.042a |
| 中氮沉降量 | 4.59±0.023a | 3.09±0.008b | 4.10±0.061b | |
| 高氮沉降量 | 4.42±0.065a | 2.63±0.032c | 4.34±0.037c | |
| 相对生长高度 | 自然沉降量 | 1.000a | 1.000a | 1.000a |
| 中氮沉降量 | 1.096 a | 0.952a | 1.445b | |
| 高氮沉降量 | 1.002 a | 0.884b | 1.641c | |
| 相对地上生物量 | 自然沉降量 | 1.000 a | 1.000 a | 1.000 a |
| 中氮沉降量 | 1.115 b | 0.995 a | 1.534 b | |
| 高氮沉降量 | 1.115 b | 0.964 b | 1.699 c | |
| 相对地下生物量 | 自然沉降量 | 1.000 a | 1.000 a | 1.000 a |
| 中氮沉降量 | 0.988 a | 0.975 a | 1.581 b | |
| 高氮沉降量 | 0.976 a | 0.944 b | 1.409 c |
注:字母相同表示差异不显著,字母不同表示差异显著。
3种稳定斑块优势物种的生长高度和生物量对氮沉降响应最强烈,且3种斑块优势物种对氮沉降量的响应不同,随着氮沉降量的增加,小叶章斑块的变化最大,牛皮杜鹃斑块的变化最小。与对照相比,中、高氮处理的牛皮杜鹃生长高度、地上生物量略高,地下生物量略低,差异不显著(P>0.05);中、高氮处理的笃斯越橘生长高度、地上生物量、地下生物量都呈现下降趋势,高氮处理与对照和中氮处理差异显著(P<0.05);随着氮沉降量的增高小叶章的生长高度、地上生物量、地下生物量指数差异显著,明显增高(P<0.05)(表1)。
随着氮沉降量的增加,不稳定斑块小叶章的多度上升;中、高氮沉降量下,轻度侵入的斑块变成中度入侵的斑块,中度侵入的斑块变成重度侵入斑块,重度侵入斑块小叶章的多度也有大幅度提高。就增幅而言,笃斯越橘斑块的小叶章的多度大于牛皮杜鹃斑块(表2)。
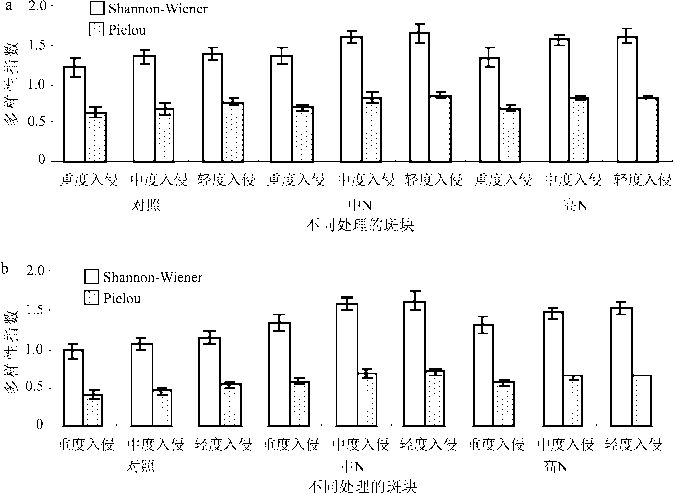
在小叶章侵入初期,草本植物的种类增多,随着小叶章的侵入加剧,牛皮杜鹃和笃斯越橘等原生植物逐步被取代。因此,轻度侵入的斑块多样性指数最高,重度侵入的斑块多样性指数最低(图1)。相同侵入程度的笃斯越橘斑块的多样性指数小于牛皮杜鹃斑块的多样性指数。随着氮沉降量的增加,不稳定斑块的物种多样性指数都在增加。中氮沉降处理的牛皮杜鹃和笃斯越橘组3种入侵程度的斑块多样性指数增加最多。
图1 牛皮杜鹃组(a)和笃斯越橘组(b)α多样性变化
Fig.1 Changes of α diversity in the group of Rhododendron chrysanthum (a) and Vaccinium uliginosum(b)
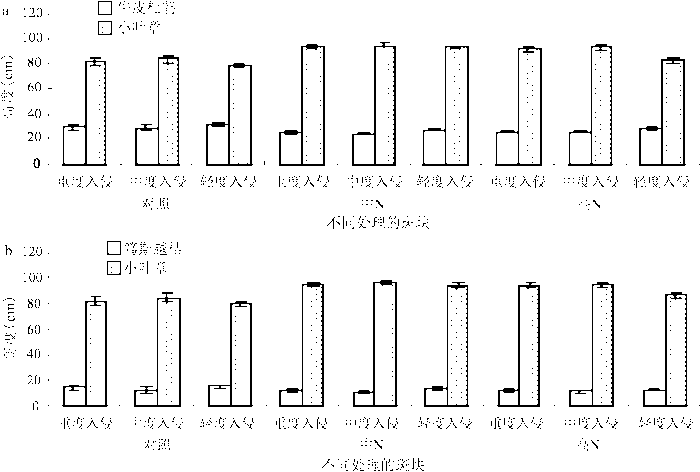
随着氮沉降量的增加,不稳定斑块中主要物种牛皮杜鹃和笃斯越橘的高度和生物量在下降,小叶章高度和生物量在上升(图2和表3)。中、高氮沉降量下笃斯越橘和牛皮杜鹃与小叶章的高度差都大于对照自然沉降量的,且中氮沉降量下的高度差最大。笃斯越橘和小叶章的高度差大于牛皮杜鹃和小叶章的高度差,随着氮沉降量的增加,这种差异在加大。随着氮沉降量的增加,小叶章生物量在斑块中占的比例越来越大,笃斯越橘组的小叶章所占生物量比例大于牛皮杜鹃组的小叶章比例,小叶章在笃斯越橘组的竞争优势更强,牛皮杜鹃群落比笃斯越橘群落抵抗小叶章侵入的能力强。
图2 不稳定斑块中牛皮杜鹃、笃斯越橘和小叶章高度对比
Fig.2 The contrast of the height of Rhododendron chrysanthum,Vaccinium uliginosum and Deyeuxia angustifolia in unstable patches
氮沉降影响的研究主要集中于沉降速率大的地区,但较小沉降速率也能对那些转化额外输入氮能力弱的生态系统造成显著影响。由于初级生产率低和土壤微生物活动弱,高山苔原对高氮沉降量产生的生物作用很敏感[21]。
通过4 a的人工模拟氮沉降量增加试验发现,长白山高山苔原带的主要优势种与入侵种对氮沉降量的响应不相同。氮沉降量增加对入侵种小叶
表2 不稳定斑块中不同氮沉降量小叶章的多度变化
Table 2 Abundance changes of Deyeuxia angustifolia in unstable patches with different nitrogen deposition
| 牛皮杜鹃组 | 笃斯越橘组 | |||||
|---|---|---|---|---|---|---|
| 重度侵入 | 中度侵入 | 轻度侵入 | 重度侵入 | 中度侵入 | 轻度侵入 | |
| 自然沉降量 | 0.77 ±0.003a | 0.52±0.002a | 0.28±0.001a | 0.79 ±0.005a | 0.55±0.002a | 0.26 ±0.001a |
| 中氮沉降量 | 0.83±0.005b | 0.76±0.004b | 0.49 ±0.003b | 0.86 ±0.006b | 0.72±0.004b | 0.61 ±0.003b |
| 高氮沉降量 | 0.83 ±0.006b | 0.75±0.003b | 0.50±0.004 | 0.93±0.009c | 0.85 ±0.007c | 0.66 ±0.005b |
表3 不稳定斑块中牛皮杜鹃、笃斯越橘和小叶章生物量之比
Table 3 The contrast of biomass of Rhododendron chrysanthum,Vaccinium uliginosum and Deyeuxia angustifolia in unstable patches
| 对照 | 中氮 | 高氮 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 重度侵入 | 中度侵入 | 轻度侵入 | 重度侵入 | 中度侵入 | 轻度侵入 | 重度侵入 | 中度侵入 | 轻度侵入 | ||
| 牛皮杜鹃组 | 地上生物量之比 | 0.86 | 4.45 | 9.70 | 0.59 | 3.39 | 8.49 | 0.38 | 2.88 | 6.39 |
| 地下生物量之比 | 2.55 | 3.23 | 4.87 | 1.97 | 2.93 | 4.35 | 1.76 | 2.58 | 3.74 | |
| 笃斯越橘组 | 地上生物量之比 | 0.46 | 2.18 | 6.29 | 0.34 | 1.76 | 4.23 | 0.23 | 1.16 | 2.20 |
| 地下生物量之比 | 2.46 | 3.04 | 5.31 | 1.64 | 2.46 | 4.31 | 1.31 | 2.12 | 3.54 | |
章生长有明显的促进作用,这一研究结果和已有的研究结论是一致的,因为小叶章是喜氮的草本植物,其生长量与丰富度会随有效氮数量的增加而增加[31~34]。
研究表明氮沉降量增加对牛皮杜鹃影响不显著,在中氮沉降情况下,对牛皮杜鹃生长略有促进,高氮沉降量对牛皮杜鹃生长有轻微抑制作用。氮沉降量增加抑制了笃斯越橘的生长,高氮沉降量比中氮沉降量对笃斯越橘的生长的抑制作用更明显。这可能是近年来长白山苔原植被出现低矮稀疏化现象的主要原因,氮沉降量增加抑制了苔原原生植物特别是笃斯越橘等灌木的生长。我们的一个关键发现是无小叶章侵入的牛皮杜鹃斑块、无小叶章侵入的笃斯越橘斑块在不同氮沉降量条件下,群落结构变化并不明显,这和瑞典北方的亚高山苔原优势种生长缓慢的小灌木被快速生长的草本代替的结论不一致[27]。这一结论也同时说明氮沉降量增加不是草本植被入侵苔原带的直接原因。但是小叶章一旦通过其它机制入侵到苔原带[39],小叶章侵入到牛皮杜鹃和笃斯越橘斑块中,就会形成小叶章与牛皮杜鹃或笃斯越橘竞争局面。我们发现随着氮沉降量的增加,不仅能促进小叶章生长,加强小叶章的竞争能力,使之逐渐取代牛皮杜鹃或笃斯越橘,成为优势种,而且促进了其他草本植物的侵入,提高了苔原带的物种多样性。可见,氮沉降量增加能强化小叶章等草本植物的侵入,推动高山苔原向高山草甸转化。这个结论能很好地解释长白山西坡于20世纪80年代末,小叶章开始侵入苔原带,进入21世纪后逐渐形成稳定的以小叶章为优势种群落的原因。小叶章上侵苔原带可能是多要素耦合的结果,对岳桦林风倒导致小叶章种子雨强度增加、苔原带侵蚀、退化导致小叶章种子易于着床等与氮沉降量增加的耦合作用,尚需进一步实验证实。
总之,随着氮沉降量的不断增加,与牛皮杜鹃、笃斯越橘等苔原带本地种相比,更有利于侵入的草本植物小叶章的生长;草本植物一旦通过某种机制入侵到苔原带后,氮沉降量增加将强化草本植物的竞争能力,逐渐取代本地的牛皮杜鹃或笃斯越橘等灌木,成为优势种,长白山苔原带将面临退化与草甸化。
The authors have declared that no competing interests exist.
| [1] |
氮沉降对森林土壤主要温室气体通量的影响 [J]. |
| [2] |
干旱半干旱区氮沉降生态效应研究进展 [J].
|
| [3] |
Plant and soil N response of southern californian semiarid shrub lands after 1 year of experimental N deposition [J].
|
| [4] |
Microbial responses to long-term N deposition in a semiarid grass land [J].
|
| [5] |
Nitrogen deposition and herbivory affect biomass production and allocation in an annual plant [J].
|
| [6] |
森林土壤氮素转换及其对氮沉降的响应 [J]. |
| [7] |
氮沉降对森林生物多样性的影响 [J]. |
| [8] |
McDowel W H ,Townsend A R,et al.The globalization of N deposition:ecosystem consequences in tropical environments [J].
|
| [9] |
Ammonia in the atmosphere: a review on emission sources, atmospheric chemistry and deposition on terrestrial bodies [J].
|
| [10] |
Alpine plant functional group responses to fertiliser addition depend on abiotic regime and community composition [J]. |
| [11] |
鼎湖山区域大气降水特征和物质元素输入对森林生态系统存在和发育的影响 [J]. |
| [12] |
太湖地区氮素湿沉降动态及生态学意义:以常熟生态站为例 [J].
|
| [13] |
Chen X Y J.Mulder Y H.Wang,et al.Atmospheric deposition, minerlization and leaching of nitrogen in subtropical forested catchments,South China [J].
|
| [14] |
大气氮沉降对阔叶林红壤淋溶水化学模拟研究 [J].
|
| [15] |
MacDonald J A,Dise N B,Matzner E,et al.Nitrogen input together with ecosystem nitrogen enrichment predict nitrate leaching from European forests [J].
|
| [16] |
Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis [J]. |
| [17] |
Estimated global nitrogen deposition using NO2 column density [J]. |
| [18] |
大气氮沉降研究进展 [J]. |
| [19] |
陆地生态系统氮沉降增加的生态效应 [J]. |
| [20] |
Constraints of nutrient availability on primary production in two alpine tundra communities [J].
|
| [21] |
Nitrogen deposition and the biodiversity of boreal forests:implications for the nitrogen critical load [J]. |
| [22] |
Response to amoniumand nitrate by a mycorrhizal annual invasive grasms and native shrub in southern California [J]. |
| [23] |
Roelofs J G M.The effects of airborne nitrogen pollutants on species diversity in natura land seminatural European vegetation [J]. |
| [24] |
Themineral nutrition of wild plants revised: a reevaluation of processes and patterns [J]. |
| [25] |
Forecasting alpine vegetation change using repeat sampling and a novel modeling approach [J]. |
| [26] |
Nitrogen critical loads for alpine vegetation and terrestrial ecosystem response: Are we there yet? [J]. |
| [27] |
Decoupled long-term effects of nutrient enrichment on aboveground and belowground properties in subalpine tundra [J]. |
| [28] |
长白山西坡小叶章侵入苔原带调查与机理分析 [J]. |
| [29] |
许嘉巍,梁宇,等.火山干扰下的长白山植被分布规律 [J]. |
| [30] |
Ecosystem response to 15 years of chronic nitrogen additions at the Harvard Forest LTER, Massachusetts, USA [J]. |
| [31] |
Effects of elevated nitrogen deposition on the field-layer vegetation in coniferous forests [J]. |
| [32] |
Nitrogen deposition and the biodiversity of boreal forests: implications for the critical load [J].
|
| [33] |
模拟氮沉降对三江平原湿地小叶章生物量及分配的影响 [J].
|
| [34] |
小叶章对氮沉降的生理生态响应 [J]. |

/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}