周勋 , 范泽孟, 岳天祥
, 范泽孟, 岳天祥
ZHOU Xun, FAN Zemeng, YUE Tianxiang
通讯作者:
收稿日期: 2016-04-25
修回日期: 2016-11-1
网络出版日期: 2017-04-20
版权声明: 2017 《地球信息科学学报》编辑部 《地球信息科学学报》编辑部 所有
基金资助:
作者简介:
作者简介:周 勋(1991-),男,安徽宁国人,硕士生,研究方向为气候变化与生态系统响应、生态模型与系统模拟。E-mail:zhoux@lreis.ac.cn
展开
摘要
针对黑河流域植被类型空间分布的垂直地带性特征,本文基于支持向量机算法构建黑河流域植被类型空间分布的模拟分析模型,并运用Kappa系数和混淆矩阵检验方法对模拟精度进行检验。验证结果显示,模型总体精度(OA)值为75.54%,Kappa系数值为0.66,表明了该方法在植被分布模拟上具有较好的结果,适用于区域尺度下植被类型分布的空间模拟。模拟结果表明,该方法对半灌木-矮半灌木荒漠和温带禾草-杂类草草甸草原类型的模拟精度最高,分别为(90.20%和90.02%);分布面积最大的植被类型(如半灌木-矮半灌木荒漠,灌木荒漠、嵩草-杂类草高寒草甸等)相比于其他面积较小的植被类型具有显著优异的模拟结果;人工经济作物、荒漠植被类型以及草原草甸等植被类型对于所选环境因子的敏感性更强,而灌丛类型和乔木类型的模拟结果在不同类型间的波动较大;空间分布上,环境要素差异性明显、植被类型丰富的上游地区具有更好的模拟结果,优于地势平坦、气候差异性小的黑河中下游地段,但模拟结果在景观形态上具有更高的破碎度。
关键词:
Abstract
According to the vertical zonality characteristic of vegetation type in Heihe River Basin, we established an analysis model of the vegetation distribution of Heihe River Basin at large scale based on Support Vector Machine algorithm. Kappa coefficient and the confusion matrix were used to validate the accuracy and performance of the model. The Overall Accuracy (OA) is 75.54% and Kappa coefficient is 0.66, indicating that this method was qualified to simulate vegetation distribution at regional scale. The results show that, semi-shrub, dwarf semi-shrub desert and temperate grasses-forbs meadow steppe have the highest simulation accuracy with OA of 90.20% and 90.02%, respectively. Vegetation types with large area such as semi-shrub and dwarf semi-shrub desert, shrub desert and Kobresia spp-forb high-cold meadows have much better accuracy than other vegetation types with small area. Artificial economic crops, desert vegetation types, and grassland and meadow are more sensitive to the chosen environmental factors. For shrub and arbor, simulation results differ among vegetation types. In the aspect of spatial distribution, upstream area with obvious distinctions in both vegetation types and environmental factors, has a better simulation results than middle and downstream area of Heihe River Basin, which are flat in terrain and have a small climate variation. Also, the simulation results of the upstream area have a higher degree of fragmentation in the landscape pattern.
Keywords:
植被类型空间分布及其结构与相应的生态环境条件密切相关[1]。植被分布与相关环境因子之间关系的研究是植被空间分布格局研究的核心内容,深入探讨其相互作用关系、产生条件、速率和作用范围,是认识植被在空间分布上的变化规律、预测未来植被分布变化趋势的前提条件,也是国家制定有关生态环境保护和规划政策的基础。植被分类是植被研究的重要组成部分,也是植被研究中最复杂的问题之一。目前,植被空间分布研究主要采用的方法包括野外样方-样线实地调查方法、遥感与GIS技术结合的方法以及空间统计模型的方法。传统上的样方-样线法,需要进行大量的野外实地调查工作来得到样点的植被信息[2],其缺点是需要耗费大量时间和劳动力,而研究区中很多地方不可达,通常情况下仅适用于小空间尺度的研究[3]。20世纪70年代以来,遥感技术和GIS技术在植物信息提取和分类等方面都得到了很大发展。高精度、高分辨率和高光谱遥感技术的发展和应用,通过监测植被结构信息和分析植被指数来进行植被类型判别,具有速度快、鲁棒性高、稳定等特点,提高了识别与分类的精度[4]。人工神经网络(ANN)、广义增强模型(GBM)、随机森林模型(RF),、分类树分析模型(CTA)、广义线性模型(GLM)和广义相加模型(GAM)等统计模型也是分析植被空间分布的重要方法,均已有广泛的应用[5]。
植被分类系统是复杂的、多层次的非线性系统,探究环境要素和植被分布相互关系的机制,建立模型是一种有效的手段[6]。支持向量机(Support Vector Machine,SVM)理论是基于结构风险最小化理论的一种机器学习算法,通过低维到高维内积空间的映射构造线性分类器。在利用SVM算法对ASTER遥感数据的分类研究中发现,SVM方法在解决小样本、非线性及高维模式识别问题中训练速度和收敛速度快,而且具有有较好的泛化能力和较高的分类精度[7-8]。另外,SVM方法在决策级多源遥感影像的融合研究中同样能取得很好的模拟精度[9-11],而且SVM方法在城市植被类型分类过程中的分类精度也明显高于传统的分类方法[12],尤其是在基于高光谱、高分辨率遥感影像进行植被分类方面[13]。但是,如何针对实地大量采样数据、结合气候、地形、人口等生境因子,对植被类型空间分布研究则涉及很少。因此,以黑河流域为研究对象,选择气候、地形、人口因子等环境因子,在对每种类型的样本进行训练确定核函数的最优参数的基础上,基于SVM方法构建植被类型分布的空间分布模型实现黑河流域植被类型空间分布的定量模拟分析。
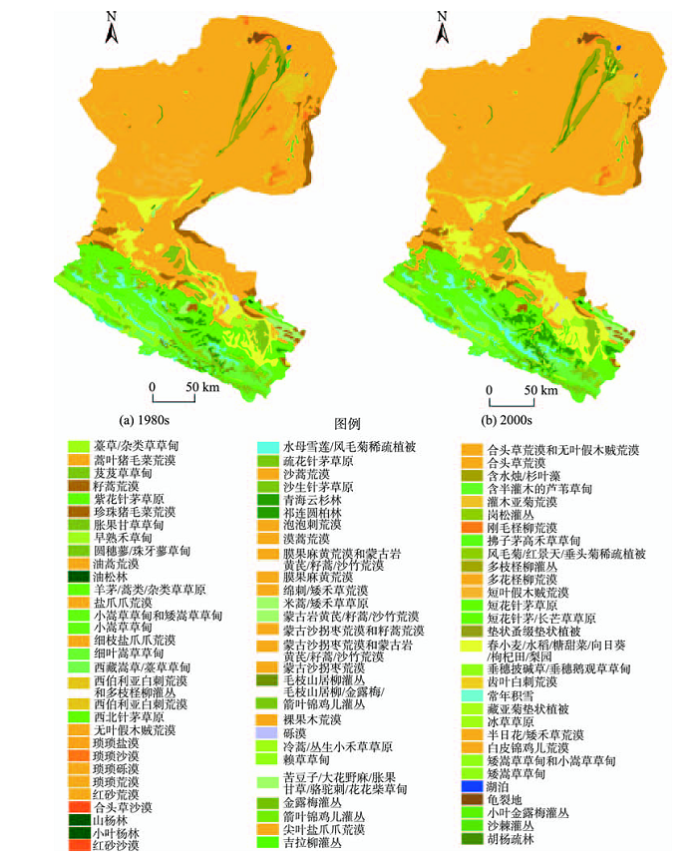
黑河发源于青藏高原东北部,干流长度超过820 km,流域总面积超过14万km2,是中国第二大内陆河。黑河流域是典型的大陆性干旱气候,从上游至下游,全流域的生物温度逐渐升高,降雨量则逐渐减少。其中,上游区段植被具有明显的垂直分布的规律性。中下游流域的蒸发率极其强烈,主要分布地带性的半灌木荒漠植被和温带小灌木植被等,而部分绿洲地区则主要以人工栽培作物和林网分布,下游的三角洲地带则主要稀疏分布有胡杨、沙枣、柽柳、白刺等荒漠植被。
DEM数据由国家基础地理信息中心发布的中国1:25万数字等高线和高程点生成,结合黑河流域20世纪80年代植被类型分布数据、2011-2013年植被样方调查数据、生态水文样带调查数据、部分多源高分辨率遥感影像数据,以及2014年野外实地样方调查数据,实现最新植被分布图中各植被群系边界的数字化,完成数据更新,得到最新的黑河流域植被类型空间分布(图1)。
图1 1980s和2000s黑河流域植被类型的空间分布对比
Fig, 1 Comparison of spatial distribution of vegetation in Heihe river Basin in 1980s and 2000s
气候变化通过改变植物种生境影响了植被分布及其生态系统过程,而气温和降水作为最主要的气候要素,其时空分布模式的变化将改变水资源和营养物质的时空分布,影响植被的光合作用、呼吸作用和土壤有机碳的分解等。地形因素主要包括海拔、地面坡度、坡向等多因子,决定了光照、风、水热条件等自然要素的空间分配,从而影响土地利用的方式及其空间分布格局。高海拔引起的气候渐变规律同时导致了植被垂直分谱的特性。土壤是植物生长的基础,为植物提供营养、水分,不同的土壤类型对应着不同的土地覆盖格局。人口快速增长情况下,尤其是在黑河流域中游的河西走廊上,人类活动植被类型及其分布变化具有很强的干扰和驱动作用,主要表现在对植被面积、景观、结构和植被质量的影响上,生态过渡带和脆弱区的环境后果更为严重,如过度放牧、过度垦荒、不节制开发等。同时,城市化进程加快,城市建设用地的增长导致了植被分布范围的缩小和转移。人口信息和交通可达性是相关规划政策和人为活动干扰对植被分布影响的侧面反映,同时流域内大量人工植被和经济作物的分布也收到极大影响,所以将人口密度、交通可达性考虑进模型参数中,从而使黑河流域中游的植被类型及分布模拟更加接近真实情况。
基于上述分析,运用黑河流域气象常规观测站点21个,周边全国站点13个的气象观测数据,提取1980-1989年,2000-2009年2个时段内记录的每日降水、气温值,整理得到各站点位置2个时段分别的10年平均值,使用高速度高精度曲面建模方法(HASM)方法模拟得到黑河流域的空间分辨率为500 m的年平均降水、平均生物温度的空间分布数据[14-15],其中平均生物温度和降水量的平均误差仅为0.45 ℃和5.20 mm。另外,利用HASM方法模拟获得的500 m分辨率的DEM、坡度坡向数据、人口分布数据、交通可达性数据、土壤质地和土壤有机质分布数据,并根据DEM数据提取了黑河流域的坡度、坡向数据。
由于黑河流域DEM数据(889~5393 m)、多年平均降水数据(8.61~854.38 mm)、多年平均生物温度数据(0.21~13.32 ℃)、人口数据(0~18 722人/km),交通可达性数据(1636~29 733 m)等生态环境参数数据的数值范围具有很大的差异性,为避免部分因子在模型构建过程中占据过多的权重,影响模型性能。因此,对基础数据机进行数据归一化处理,从而解决数据指标之间的可比性。
数据归一化处理方法如式(1)所示。
式中:
植被类型的空间分布具有典型的空间复杂性和非线性特征,运用线性分类方法无法满足。而支持向量机方法(SVM)则能为解决非线性回归的求解问题提供了一种新的新思路。相比传统的方法,其在速度和精度上都有明显的优势,在有限样本情况下的学习精度与学习能力都得到了最佳折衷,具有较好的推广能力。支持向量机方法是一种建立在统计学理论和基于结构风险最小化原则基础上的机器学习系统,经过了严密的理论证明,即使在样本量较少的情况下,同样能获得良好的分类结果和统计规律。通常适用的分析基于数据线性可分的情况下,但当遇到线性不可分的案例时,SVM会通过非线性的映射算法把原本线性不可分的样本数据从低维输入空间转到高维特征空间,在高维特征空间中寻得最优分类超平面,得到了全局最优分类器,同时以某个特定的概率使分类器在整个样本空间的期望风险满足一定的上界。支持向量指距离低维空间最优分类线或高维空间的最优分类面最近的所有向量。运用SVM方法构建植被类型空间分布模拟模型的原理及方法如下。
核函数是一个函数K,对于所有的x,z,满足(是从输入空间X到特征空间F的映射),常用核函数的类型主要有4个:① 线性核函数:
同理,构造了Lagrange函数(式(3))。
式中:
最终获得非线性回归函数(式(6))。
构建植被类型空间分布模拟模型的关键则是实现对SVM参数的C和
运用网格搜索法进行最优核参数的选择,首 先设定SVM参数σ和惩罚系数C的范围,在
在对数据集进行归一化的基础上,以每个栅格格点所包含的各环境要素数据值构成向量作为样本数据,采用不放回的随机采样方法,按照固定的采样比例获得训练样本数据集,用于确定模型中的参数并构建模型,剩余数据集用于测试分类效果。每组植被类型使用的训练数据集和测试数据集都采用了统一映射函数来进行处理,保证训练模型和测试模型都使用同样的标准。样本的选择对于植被分布模拟结果至关重要,故要具备全面性与典型性。研究中以新完成的黑河流域2000 s植被分布数据为基础数据,分别提取出每种植被类型的分布栅格数据,共具有514 450个栅格点,在训练样本获取时,分别对包含植被分布的斑块和空白区域采用随机采样方法取5%的样点,共得到25 722个向 量单元作为模型的训练数据集。使用训练数据集使用SVM方法进行训练,使用网格搜索方法确定模型中的最优[C,σ]参数,并使用K-折交叉验证法(K-fold Cross Validation)计算该参数下模拟的准确率,挑选出最优的参数对,测试模型的性能。数据中其他未作为训练样本的数据栅格单元作为模拟数据集完成模型的构建,运用总体精度(OA)、Kappa系数方法对模拟结果的精度进行了验证分析。由于不同植被类型对于环境因子敏感性的差异显著,因此根据不同的植被类型得到不同的模型最优化参数,如表1所示。
表1 最优参数列表及测试结果
Tab. 1 The optimal parameters and testing results
| 编号 | 植被类型 | C | gamma | 准确率 | kappa | 植被总面积/hm2 |
|---|---|---|---|---|---|---|
| 1 | 一年一熟粮食作物及耐寒经济作物田、落叶果树园 | 32 | 16 | 0.97 | 0.76 | 84.03 |
| 2 | 亚热带、热带常绿阔叶、落叶阔叶灌丛 | 128 | 256 | 0.99 | 0.27 | 0.09 |
| 3 | 亚高山落叶阔叶灌丛 | 4 | 128 | 0.98 | 0.35 | 28.58 |
| 4 | 温带针叶林 | 128 | 32 | 0.99 | 0.15 | 0.26 |
| 5 | 温带落叶小叶林 | 32 | 10 | 0.99 | 0.15 | 7.79 |
| 6 | 温带落叶阔叶林 | 16 | 10 | 0.99 | 0.08 | 0.11 |
| 7 | 温带落叶灌丛 | 64 | 256 | 0.99 | 0.61 | 25.18 |
| 8 | 温带禾草、杂类草草甸草原 | 64 | 1 | 0.99 | 0.00 | 0.34 |
| 9 | 温带丛生禾草草原 | 64 | 4 | 0.98 | 0.67 | 52.19 |
| 10 | 温带丛生矮禾草、矮半灌木荒漠草原 | 8 | 128 | 0.99 | 0.64 | 25.80 |
| 11 | 嵩草、杂类草高寒草甸 | 1 | 32 | 0.95 | 0.69 | 107.96 |
| 12 | 禾草、杂类草盐生草甸 | 16 | 256 | 0.99 | 0.55 | 11.12 |
| 13 | 禾草、杂类草草甸 | 8 | 100 | 0.99 | 0.41 | 1.82 |
| 14 | 禾草、薹草高寒草原 | 8 | 64 | 0.98 | 0.39 | 21.53 |
| 15 | 寒温带和温带山地针叶林 | 8 | 64 | 0.98 | 0.45 | 21.77 |
| 16 | 灌木荒漠 | 4 | 256 | 0.91 | 0.60 | 186.47 |
| 17 | 高山稀疏植被 | 32 | 10 | 0.98 | 0.39 | 27.09 |
| 18 | 高山垫状植被 | 16 | 256 | 0.99 | 0.18 | 3.24 |
| 19 | 多汁盐生矮半灌木荒漠 | 8 | 128 | 0.98 | 0.59 | 33.74 |
| 20 | 草原化灌木荒漠 | 128 | 256 | 0.99 | 0.36 | 0.90 |
| 21 | 草原 | 32 | 64 | 0.99 | 0.79 | 3.54 |
| 22 | 半灌木、矮半灌木荒漠 | 4 | 128 | 0.88 | 0.71 | 570.79 |
| 23 | 矮半乔木荒漠 | 8 | 256 | 0.97 | 0.55 | 60.49 |
| 24 | 湖泊水体等无植被地段 | 16 | 1 | 0.99 | 0.28 | 7.82 |
由于不同植被类型对于环境因子的敏感性不同,模型构建中最优[C,σ]参数的差异性很明显,因此如果按照同一个最优参数标准对所有植被类型进行整体分类的误差较大,而根据不同植被类型建立多个空间分布模型进行模拟,其结果更有特征性和合理性。
从准确率上看,除去半灌木-矮半灌木荒漠植被类型的85.63%外,其余23种类型的准确率均超过了90%,其中亚热带-热带常绿阔叶-落叶阔叶灌丛、温带针叶林、温带落叶阔叶林、温带禾草-杂类草草甸草原等植被类型超过了99%;Kappa系数上来看,最高的是草原类型,其Kappa值达到了0.79,其次是一年一熟粮食作物及耐寒经济作物田-落叶果树园和半灌木-矮半灌木荒漠类型,分别达到了0.76和0.71。综合以上2项指标,草原、一年一熟粮食作物及耐寒经济作物田-落叶果树园的模拟结果最佳,准确率和Kappa系数均超过97%和0.75。
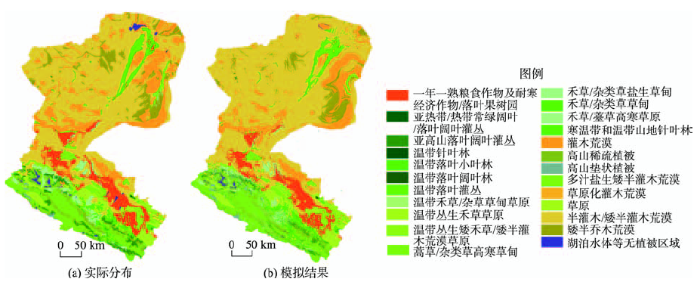
在模型运行过程中,依次提取每种植被类型的分布信息,与环境要素数据叠加,分别构建模型且完成分布模拟,对输出结果进行叠加,提取出所有重合的点,保留其他栅格点。对各重合的栅格点,进行重新构建多分类模型并完成模拟,最终将所有的模拟结果进行整合,得到完整的分布。模拟结果如图2所示,整体上来看实际植被分布数据(图2(a))和模拟结果(图2(b))具有较高的重合度,斑块结构和形状基本相似,但是模拟结果的破碎化程度更高。上游和中游的模拟效果较好,其中上游流域植被类型众多,生态系统结构复杂,包括多种乔木、灌丛、草原草甸、高山植被类型。人工经济作物主要分布在上游的东北角和中游中部,此外中游区域还有少量草甸植被类型和荒漠植被类型。下游以灌木荒漠植被类型为主,其西部和北部的模拟结果一般使用混淆矩阵进行验证得到结果如表2所示,总体精度(OA)达到了75.54%,Kappa系数为0.66;对混淆矩阵统计后整理得到表3。
图2 植被分布数据与模拟结果对比
Fig. 2 Comparison of the original vegetation distribution and the simulation results
表2 植被分布模拟结果的混淆矩阵(km2)
Tab.2 Confusion matrix of the simulation results of vegetation distribution (km2)
| A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | 合计 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | 62.54 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.11 | 0.00 | 0.60 | 2.36 | 0.04 | 0.69 | 0.12 | 0.04 | 0.04 | 1.13 | 0.00 | 0.00 | 0.97 | 0.00 | 0.23 | 3.84 | 0.00 | 0.71 | 73.39 |
| B | 0.00 | 0.06 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.25 | 0.02 | 0.00 | 0.32 |
| C | 0.00 | 0.00 | 9.18 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 1.64 | 0.03 | 3.75 | 0.01 | 0.01 | 1.99 | 0.85 | 0.05 | 1.42 | 0.00 | 0.04 | 0.00 | 0.00 | 0.62 | 0.00 | 0.01 | 19.59 |
| D | 0.01 | 0.00 | 0.00 | 0.21 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.02 | 0.01 | 0.00 | 0.30 |
| E | 0.12 | 0.00 | 0.00 | 0.00 | 6.45 | 0.00 | 0.21 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.02 | 0.01 | 0.00 | 8.14 |
| F | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.02 |
| G | 0.82 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 14.49 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.06 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 4.09 | 0.09 | 0.03 | 21.57 |
| H | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.28 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.34 |
| I | 1.30 | 0.00 | 1.22 | 0.00 | 0.57 | 0.01 | 0.00 | 0.00 | 35.38 | 1.60 | 2.15 | 0.47 | 0.06 | 0.85 | 2.06 | 0.05 | 0.10 | 0.00 | 1.18 | 0.00 | 0.02 | 2.35 | 0.00 | 0.00 | 48.77 |
| J | 3.65 | 0.00 | 0.06 | 0.00 | 0.00 | 0.03 | 0.00 | 0.00 | 1.81 | 17.86 | 0.22 | 0.02 | 0.05 | 0.84 | 0.04 | 0.07 | 0.00 | 0.00 | 0.54 | 0.00 | 0.29 | 2.24 | 0.00 | 0.00 | 27.70 |
| K | 0.04 | 0.00 | 12.12 | 0.00 | 0.00 | 0.02 | 0.10 | 0.00 | 4.03 | 0.20 | 85.23 | 0.08 | 0.32 | 5.30 | 1.77 | 0.52 | 11.24 | 0.00 | 0.00 | 0.00 | 0.00 | 0.58 | 0.00 | 0.49 | 122.02 |
| L | 1.07 | 0.00 | 0.00 | 0.02 | 0.00 | 0.00 | 0.00 | 0.04 | 0.15 | 0.05 | 0.00 | 5.46 | 0.01 | 0.00 | 0.02 | 0.08 | 0.00 | 0.00 | 0.32 | 0.00 | 0.00 | 1.75 | 0.00 | 0.00 | 8.90 |
| M | 0.07 | 0.00 | 0.04 | 0.00 | 0.54 | 0.04 | 0.00 | 0.00 | 0.02 | 0.01 | 0.20 | 0.01 | 0.71 | 0.05 | 0.21 | 0.00 | 0.02 | 0.00 | 0.00 | 0.00 | 0.00 | 0.11 | 0.00 | 0.00 | 1.48 |
| N | 0.04 | 0.00 | 2.36 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.32 | 0.64 | 4.04 | 0.00 | 0.06 | 10.06 | 1.40 | 0.01 | 1.13 | 0.00 | 0.05 | 0.00 | 0.01 | 0.83 | 0.00 | 0.02 | 21.96 |
| O | 0.02 | 0.00 | 1.35 | 0.02 | 0.00 | 0.01 | 0.01 | 0.02 | 3.76 | 0.25 | 2.31 | 0.13 | 0.31 | 0.76 | 14.97 | 0.00 | 0.21 | 0.00 | 0.17 | 0.00 | 0.05 | 0.54 | 0.00 | 0.00 | 24.82 |
| P | 2.16 | 0.00 | 0.02 | 0.00 | 0.20 | 0.00 | 0.17 | 0.00 | 0.05 | 0.16 | 0.10 | 0.84 | 0.00 | 0.01 | 0.00 | 106.30 | 0.02 | 0.00 | 5.35 | 0.00 | 0.00 | 24.70 | 5.71 | 1.45 | 147.04 |
| Q | 0.00 | 0.00 | 1.62 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.26 | 0.00 | 8.59 | 0.00 | 0.03 | 0.77 | 0.10 | 0.05 | 12.09 | 0.00 | 0.00 | 0.00 | 0.00 | 0.05 | 0.00 | 1.31 | 24.85 |
| R | 0.00 | 0.00 | 0.10 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.19 | 0.00 | 0.64 | 0.00 | 0.00 | 0.07 | 0.00 | 0.01 | 0.08 | 3.24 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | 4.33 |
| S | 1.58 | 0.00 | 0.03 | 0.00 | 0.24 | 0.00 | 0.00 | 0.00 | 1.26 | 0.80 | 0.00 | 0.04 | 0.00 | 0.15 | 0.06 | 2.20 | 0.00 | 0.00 | 18.47 | 0.00 | 0.00 | 3.05 | 0.22 | 0.01 | 27.85 |
| T | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.04 | 0.00 | 0.07 | 0.02 | 0.00 | 0.13 |
| U | 0.46 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.21 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.93 | 0.00 | 0.00 | 0.06 | 3.66 |
| V | 10.18 | 0.03 | 0.48 | 0.00 | 0.00 | 0.01 | 10.08 | 0.00 | 1.73 | 1.62 | 0.41 | 2.99 | 0.14 | 0.63 | 0.27 | 66.31 | 0.03 | 0.00 | 6.16 | 0.61 | 0.01 | 514.35 | 22.76 | 1.68 | 640.47 |
| W | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.37 | 0.00 | 0.00 | 0.00 | 6.95 | 0.00 | 0.00 | 0.53 | 0.25 | 0.00 | 10.81 | 31.47 | 0.00 | 50.38 |
| X | 0.00 | 0.00 | 0.02 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.28 | 0.00 | 0.00 | 0.04 | 0.00 | 0.18 | 0.77 | 0.00 | 0.00 | 0.00 | 0.00 | 0.02 | 0.01 | 2.05 | 3.35 |
| 合计 | 84.03 | 0.09 | 28.58 | 0.26 | 7.79 | 0.11 | 25.18 | 0.34 | 52.18 | 25.80 | 107.95 | 11.12 | 1.82 | 21.53 | 21.77 | 185.97 | 27.09 | 3.24 | 33.74 | 0.90 | 3.54 | 570.26 | 60.31 | 7.82 | 1281.38 |
| 总体精度(OA)= 75.54% Kappa系数=0.66 | |||||||||||||||||||||||||
表3 混淆矩阵中的植被类型对应表
Tab.3 Vegetation types in confusion matrix
| 编号 | 植被类型 | 植被总面积/万hm2 | 模拟结果面积/万hm2 | 精度/% |
|---|---|---|---|---|
| A | 一年一熟粮食作物及耐寒经济作物田、落叶果树园 | 84.03 | 73.39 | 74.42 |
| B | 亚热带、热带常绿阔叶、落叶阔叶灌丛 | 0.09 | 0.32 | 62.86 |
| C | 亚高山落叶阔叶灌丛 | 28.58 | 19.59 | 32.13 |
| D | 温带针叶林 | 0.26 | 0.30 | 83.00 |
| E | 温带落叶小叶林 | 7.79 | 8.14 | 78.00 |
| F | 温带落叶阔叶林 | 0.11 | 0.02 | 4.55 |
| G | 温带落叶灌丛 | 25.18 | 21.57 | 57.55 |
| H | 温带禾草、杂类草草甸草原 | 0.34 | 0.34 | 90.02 |
| I | 温带丛生禾草草原 | 52.19 | 48.77 | 67.80 |
| J | 温带丛生矮禾草、矮半灌木荒漠草原 | 25.80 | 27.70 | 69.22 |
| K | 嵩草、杂类草高寒草甸 | 107.96 | 122.02 | 78.95 |
| L | 禾草、杂类草盐生草甸 | 11.12 | 8.90 | 49.13 |
| M | 禾草、杂类草草甸 | 1.82 | 1.48 | 39.12 |
| N | 禾草、薹草高寒草原 | 21.53 | 21.96 | 46.73 |
| O | 寒温带和温带山地针叶林 | 21.77 | 24.82 | 68.78 |
| P | 灌木荒漠 | 186.47 | 147.04 | 57.16 |
| Q | 高山稀疏植被 | 27.09 | 24.85 | 44.63 |
| R | 高山垫状植被 | 3.24 | 4.33 | 89.32 |
| S | 多汁盐生矮半灌木荒漠 | 33.74 | 27.85 | 54.72 |
| T | 草原化灌木荒漠 | 0.90 | 0.13 | 4.75 |
| U | 草原 | 3.54 | 3.66 | 82.70 |
| V | 半灌木、矮半灌木荒漠 | 570.79 | 640.47 | 90.20 |
| W | 矮半乔木荒漠 | 60.49 | 50.38 | 52.18 |
| X | 湖泊水体等无植被地段 | 7.82 | 3.35 | 26.21 |
由表2、3分析表明,制图精度最高的植被类型是半灌木-矮半灌木荒漠、温带禾草-杂类草草甸草原、高山垫状植被,分别达到了90.20%、90.02%和89.32%,而温带落叶阔叶林、草原化灌木荒漠的制图精度依次仅为4.55%、4.75%,模拟结果精度低。其中温带落叶阔叶林的优势种为山杨林和小叶杨林,主要错分为禾草-杂类草草甸(31.82%)、温带丛生矮禾草-矮半灌木荒漠草原(25.00%)和嵩草-杂类草高寒草甸(20.45%);草原化灌木荒漠的优势种为半日花-矮禾草和锦刺-矮禾草,主要错分类型为半灌木-矮半灌木荒漠(67.60%)和矮半乔木荒漠(27.65%);湖泊水体等无植被地段的模拟精度为26.21%,仅高于上述两种植被类型,表明研究区内湖泊水体的分布与人文、环境因子分布的相关性程度不高。温带禾草-杂类草草甸草原的用户精度达到了88.53%,温带针叶林为85.61%,为精度最高的两种植被类型。亚热带、热带常绿阔叶、落叶阔叶灌丛植被类型的精度仅为17.05%,与其他类型具有较大差距。之后依次为草原化灌木荒漠(32.08%)和温带落叶阔叶林(33.33%)。
模拟结果与原植被分布数据对比,面积上出现了不同程度的增减。模拟后面积增加率最高的是亚热带-热带常绿阔叶-落叶阔叶灌丛类型,达到了258.33%,其模拟前总面积为0.09万hm2,对应的原植被类型应为半灌木-矮半灌木荒漠,错误模拟 面积为0.25万hm2。其依是高山垫状植被和温带针叶林,模拟前面积和增加率分别为3.24万hm2、33.64%和0.26万hm2、16.35%。错误模拟为高山垫状植被区域原对应的正确植被类型主要为嵩草-杂类草高寒草甸(0.64万hm2)和温带丛生禾草草原 (0.19万hm2);错误模拟为温带针叶林类型区域对应的正确植被类型中,面积最多的为半灌木-矮半灌木荒漠的0.02万hm2。模拟后面积减少率最高的是温带落叶阔叶林(-86.36%)和草原化灌木荒漠 (-85.28%),原分布数据中的面积分别为0.11万hm2和0.90万hm2。主要分别错误模拟为禾草-杂类草草甸(0.035万hm2)、温带丛生矮禾草-矮半灌木荒漠草原(0.028万hm2)和嵩草-杂类草高寒草甸(0.03万hm2);草原化灌木荒漠的优势种为半日花-矮禾草和锦刺-矮禾草,主要错分类型为半灌木-矮半灌木荒漠(0.61万hm2)和矮半乔木荒漠(0.25万hm2)。
通过分析发现不同的植被分布面积和类型,模拟精度存在一定的差异性。从植被类型分布面积上看,分布面积较大的植被类型具有显著优异的模拟结果,半灌木-矮半灌木荒漠(570.79万hm2)的总体精度达到了90.20%,而嵩草-杂类草高寒草甸(107.96万hm2)、一年一熟粮食作物及耐寒经济作物田-落叶果树园(84.03万hm2)对应的2种精度分别达到了78.95%、74.42%。分布面积很小的植被类型,如亚热带-热带常绿阔叶-落叶阔叶灌丛(0.09万hm2)、温带落叶阔叶林(0.11万hm2)、草原化灌木荒漠(0.90万hm2)、禾草-杂类草草甸(1.82万hm2)等,其模拟精度低于平均值。但如温带针叶林(0.26万hm2)、温带禾草-杂类草草甸草原(0.34万hm2)、草原(3.54万hm2)、高山垫状植被(3.24万hm2)等具有较好的模拟结果,精度大多接近或超过80%,其中温带禾草-杂类草草甸草原的制图精度达到了90.02%。结果表明,使用该方法对大范围分布的植被类型与环境要素间相关关系的规律性的探究效果更好,受其他生境特征接近的植被类型分布或异常数据的影响较小。分布面积极小的植被类型在分布模拟时易受到其他类型的干扰。
本文在对黑河流域生态环境参数进行采集及预处理的基础上,基于SVM方法构建了黑河流域植被类型空间分布的模拟模型,并在植被群组层次上对每种植被类型的空间分布进行了模拟,模拟结果的总体精度(OA)为75.54%,Kappa系数为0.66。精度最高的植被类型是半灌木-矮半灌木荒漠、温带禾草-杂类草草甸草原、高山垫状植被,分别达到了90.20%、90.02%和89.32%。这表明该方法在植被分布模拟上具有较好的结果,适用于区域尺度下植被类型分布的空间模拟。
在模拟分析得过程中发现,SVM方法对一年一熟粮食作物及耐寒经济作物田-落叶果树园代等人工经济作物的模拟精度(74.42%)整体高于其他植被类型的模拟分类精度。另外,主要分布在黑河上游高海拔段高寒区域的高山植被类型与环境要素显著相关。草原和草甸类型的整体模拟结果较好,其中温带和寒带草原草甸类型优于荒漠草原类型,乔木类型中针叶林模拟精度高于阔叶林类型。灌木、灌丛和荒漠植被的模拟情况,在不同类型间差异较大,如半灌木-矮半灌木荒漠类型的精度达到了90.20%,而多汁盐生矮半灌木荒漠的模拟精度仅为54.72%。
同时,黑河上游和中游地区植被类型与生境要素的相关性明显高于下游地区,这表明地形起伏度的复杂程度使得植被类型的空间分布与生境要素之间的相关性更为明显。例如,主要分布在上游东北部和中游的中部区域的乔木、草原草甸、灌丛、高山植被类型等植被类型的模拟结果明显优于地势平坦、气候差异性小的黑河下游地段。上游植被分布密集,斑块面积小、破碎程度高,整体结构上错综复杂,使模拟结果在景观形态上的破碎程度更高。
此外,本文构建的模型模拟方法尚处于初步阶段,需要在未来的研究工作中进一步提高模型参数的分辨率,更进一步优化模型参数,从而在对模型方法进行修正和完善的基础上,运用模型对植被类型空间分布的未来情景进行模拟分析,从而提升模型的适用深度和广度。
The authors have declared that no competing interests exist.
| [1] |
Predictive habitat distribution models in ecology [J].https://doi.org/10.1016/S0304-3800(00)00354-9 URL [本文引用: 1] 摘要
With the rise of new powerful statistical techniques and GIS tools, the development of predictive habitat distribution models has rapidly increased in ecology. Such models are static and probabilistic in nature, since they statistically relate the geographical distribution of species or communities to their present environment. A wide array of models has been developed to cover aspects as diverse as biogeography, conservation biology, climate change research, and habitat or species management. In this paper, we present a review of predictive habitat distribution modeling. The variety of statistical techniques used is growing. Ordinary multiple regression and its generalized form (GLM) are very popular and are often used for modeling species distributions. Other methods include neural networks, ordination and classification methods, Bayesian models, locally weighted approaches (e.g. GAM), environmental envelopes or even combinations of these models. The selection of an appropriate method should not depend solely on statistical considerations. Some models are better suited to reflect theoretical findings on the shape and nature of the species response (or realized niche). Conceptual considerations include e.g. the trade-off between optimizing accuracy versus optimizing generality. In the field of static distribution modeling, the latter is mostly related to selecting appropriate predictor variables and to designing an appropriate procedure for model selection. New methods, including threshold-independent measures (e.g. receiver operating characteristic (ROC)-plots) and resampling techniques (e.g. bootstrap, cross-validation) have been introduced in ecology for testing the accuracy of predictive models. The choice of an evaluation measure should be driven primarily by the goals of the study. This may possibly lead to the attribution of different weights to the various types of prediction errors (e.g. omission, commission or confusion). Testing the model in a wider range of situations (in space and time) will permit one to define the range of applications for which the model predictions are suitable. In turn, the qualification of the model depends primarily on the goals of the study that define the qualification criteria and on the usability of the model, rather than on statistics alone.
|
| [2] |
典型陆地生态系统对气候变化响应的定量研究 [J].https://doi.org/10.1360/972013-261 Magsci [本文引用: 1] 摘要
<p>森林、农田和草地生态系统对气候变化响应定量研究进展的综合分析表明:近一个世纪以来的森林、农田和草地生态系统对气候变化有很强的响应,诸多生态系统类型的组成、结构和分布已发生了显著变化;由于病虫害、极端气候频发,植物物种的死亡率增加、生产力出现下降趋势;典型陆地生态系统对气候响应的未来情景分析结果表明,高海拔地区和高纬度地区的生态系统类型的结构、分布、物种和生产力将发生较大变化;然而,由于生物群落的相互作用,各种生态系统对气候变化的响应很复杂,目前人类对典型陆地生态系统变化的认识仍然处在很初级的阶段. 尤其是在气候变化对植物物种的影响、干旱和极端事件的后果以及病虫害的影响等方面,还没有明确的结论;根据目前的研究积累,还无法给出气候变化对典型陆地生态系统影响的综合定量评估,需要改进区域气候模拟,尤其是降雨量的模拟,需要提高植物物种对气候、病虫害和大气成分响应的认识. 以典型陆地生态系统对全球气候变化响应机理研究成果为基础、集成空间对地观测数据和地面实测数据的多尺度生态系统动态模拟分析平台,到目前为止,仍是亟待填补的空白.</p>
A review of responses of typical terrestrial ecosystems to climate change [J].https://doi.org/10.1360/972013-261 Magsci [本文引用: 1] 摘要
<p>森林、农田和草地生态系统对气候变化响应定量研究进展的综合分析表明:近一个世纪以来的森林、农田和草地生态系统对气候变化有很强的响应,诸多生态系统类型的组成、结构和分布已发生了显著变化;由于病虫害、极端气候频发,植物物种的死亡率增加、生产力出现下降趋势;典型陆地生态系统对气候响应的未来情景分析结果表明,高海拔地区和高纬度地区的生态系统类型的结构、分布、物种和生产力将发生较大变化;然而,由于生物群落的相互作用,各种生态系统对气候变化的响应很复杂,目前人类对典型陆地生态系统变化的认识仍然处在很初级的阶段. 尤其是在气候变化对植物物种的影响、干旱和极端事件的后果以及病虫害的影响等方面,还没有明确的结论;根据目前的研究积累,还无法给出气候变化对典型陆地生态系统影响的综合定量评估,需要改进区域气候模拟,尤其是降雨量的模拟,需要提高植物物种对气候、病虫害和大气成分响应的认识. 以典型陆地生态系统对全球气候变化响应机理研究成果为基础、集成空间对地观测数据和地面实测数据的多尺度生态系统动态模拟分析平台,到目前为止,仍是亟待填补的空白.</p>
|
| [3] |
Multispectral and hyperspectral remote sensing for identification and mapping of wetland vegetation: a review [J].https://doi.org/10.1007/s11273-009-9169-z URL [本文引用: 1] 摘要
Abstract Wetland vegetation plays a key role in the ecological functions of wetland environments. Remote sensing techniques offer timely, up-to-date, and relatively accurate information for sustainable and effective management of wetland vegetation. This article provides an overview on the status of remote sensing applications in discriminating and mapping wetland vegetation, and estimating some of the biochemical and biophysical parameters of wetland vegetation. Research needs for successful applications of remote sensing in wetland vegetation mapping and the major challenges are also discussed. The review focuses on providing fundamental information relating to the spectral characteristics of wetland vegetation, discriminating wetland vegetation using broad- and narrow-bands, as well as estimating water content, biomass, and leaf area index. It can be concluded that the remote sensing of wetland vegetation has some particular challenges that require careful consideration in order to obtain successful results. These include an in-depth understanding of the factors affecting the interaction between electromagnetic radiation and wetland vegetation in a particular environment, selecting appropriate spatial and spectral resolution as well as suitable processing techniques for extracting spectral information of wetland vegetation. KeywordsBiomass-Leaf area index-Mapping-Remote sensing-Water content-Wetland vegetation
|
| [4] |
遥感植被分类方法的概述及发展 [J].
卫星遥感技术的发展为大范围作物的识别分类提供了新的方法,90年代以来国外主要使用卫星遥感结合植物表面温度、地形等非遥感要素对地面植被进行判识分类,我国自80年代以来应用图像纹理、植被指数变化、植被指数的动态聚类及经验正交函数等方法对大范围植被分类进行了研究.近年来,为提高植被分类精度已在提高资料的空间分辨率、利用微波遥感,综合应用"3S"技术等方面做了努力.
Review of vegetation classification based on remote sensing method [J].
卫星遥感技术的发展为大范围作物的识别分类提供了新的方法,90年代以来国外主要使用卫星遥感结合植物表面温度、地形等非遥感要素对地面植被进行判识分类,我国自80年代以来应用图像纹理、植被指数变化、植被指数的动态聚类及经验正交函数等方法对大范围植被分类进行了研究.近年来,为提高植被分类精度已在提高资料的空间分辨率、利用微波遥感,综合应用"3S"技术等方面做了努力.
|
| [5] |
延河流域典型物种分布预测模型比较研究 [J].
物种分布预测一直以来都是生态学研究的重要内容之一。应用生态学的发展为物种分布预测提供了众多强有力的模型,在推进物种分布预测进展的同时,也增加了合适模型选择的难度。评价和比较不同模型的预测效果,对于模型的选择和应用具有非常重要的意义。以黄土丘陵区延河流域为研究区,采用R语言和BIOMOD程序包为平台,选择人工神经网络(artificial neural networks,ANN)等9个较常用的物种分布模型,比较它们在物种分布预测精度上的差异,为物种分布预测模型的选择提供依据,也为进一步预测未来气候变化情景下物种空间分布的变化奠定基础。研究结果表明,不同模型对不同物种的模拟精度差异明显。根据Kappa,TSS和Roc评价方法,9个模型对百里香(Thymus mongolicus)分布的预测精度最高;对铁杆蒿(Artemisia gmelinii)分布的模拟精度最差;而对其余物种分布的模拟精度均比较理想,其中以随机树RF模型最好。
Comparison of predictive models for representative species distribution in Yanhe river basin.
物种分布预测一直以来都是生态学研究的重要内容之一。应用生态学的发展为物种分布预测提供了众多强有力的模型,在推进物种分布预测进展的同时,也增加了合适模型选择的难度。评价和比较不同模型的预测效果,对于模型的选择和应用具有非常重要的意义。以黄土丘陵区延河流域为研究区,采用R语言和BIOMOD程序包为平台,选择人工神经网络(artificial neural networks,ANN)等9个较常用的物种分布模型,比较它们在物种分布预测精度上的差异,为物种分布预测模型的选择提供依据,也为进一步预测未来气候变化情景下物种空间分布的变化奠定基础。研究结果表明,不同模型对不同物种的模拟精度差异明显。根据Kappa,TSS和Roc评价方法,9个模型对百里香(Thymus mongolicus)分布的预测精度最高;对铁杆蒿(Artemisia gmelinii)分布的模拟精度最差;而对其余物种分布的模拟精度均比较理想,其中以随机树RF模型最好。
|
| [6] |
气候变化对植物及植被分布的影响研究进展 [J].https://doi.org/10.3969/j.issn.1003-6504.2009.06.020 URL [本文引用: 1] 摘要
准确预测气候变化对植物及植被分布的影响趋势对于科学认识气候变 化对生物多样性的影响和对策制定具有重要的意义.近年来,气候变化对植物及植被分布影响研究在我国广泛展开.文章对这些研究所采用的气候情景、预测方法和 气候变化等对植物及植被分布的影响趋势进行了总结分析,对存在的问题进行了讨论,对未来的趋势提出了若干展望.总体上,目前所用气候情景比较单一、不同预 测方法比较研究不够,对土壤和生物因素考虑不充分、涉及植物种类太少.气候变化将使一些植物分布范围减少、甚至消失,而使另一些植物分布范围则会扩大.未 来研究应加强气候情景改进和不同方法的比较研究,充分考虑土壤和生物等因子、加强预测方法检验,同时开展植物物种多样性和丰富度对气候变化响应方面的研 究.
Advances in the effects of climate change on the distribution of plant species and vegetation in China. https://doi.org/10.3969/j.issn.1003-6504.2009.06.020 URL [本文引用: 1] 摘要
准确预测气候变化对植物及植被分布的影响趋势对于科学认识气候变 化对生物多样性的影响和对策制定具有重要的意义.近年来,气候变化对植物及植被分布影响研究在我国广泛展开.文章对这些研究所采用的气候情景、预测方法和 气候变化等对植物及植被分布的影响趋势进行了总结分析,对存在的问题进行了讨论,对未来的趋势提出了若干展望.总体上,目前所用气候情景比较单一、不同预 测方法比较研究不够,对土壤和生物因素考虑不充分、涉及植物种类太少.气候变化将使一些植物分布范围减少、甚至消失,而使另一些植物分布范围则会扩大.未 来研究应加强气候情景改进和不同方法的比较研究,充分考虑土壤和生物等因子、加强预测方法检验,同时开展植物物种多样性和丰富度对气候变化响应方面的研 究.
|
| [7] |
Classification using ASTER data and SVM algorithms: The case study of Beer Sheva, Israel [J]. |
| [8] |
SVM texture classification for tropical vegetation mapping [J].https://doi.org/10.1117/12.977182 URL [本文引用: 1] 摘要
Nowadays, remote sensing is an essential science in French Polynesia because of its extended territory and the remoteness of its 120 islands. There is a strong need to study the vegetation cover and its evolution (biodiversity threat, invasive species, etc.). A growing satellite images database has been acquired throughout, giving access to very high resolution optical images such as Quickbird data. These data allow accessing the vegetation canopy spectral and contextual information, texture classification has proved to be an efficient tool to map the complex vegetation found in tropical regions. The main goal of this paper is to propose an optimized SVM multispectral-texture classification method for tropical vegetation mapping. One of the texture computation drawbacks is the window treatment size, which is related to the largest texture element size. In complex tropical vegetation cover, this parameter leads to very small ground truth learning database, inducing a significant degradation of the classifications accuracy. We propose to increase the thumbnail numbers using an under-sampling method, optimizing the size and the number of the thumbnails. The other drawback is the high dimensionality of the problem when dealing with multispectral textures. We thus propose to rank and select the most pertinent textures attributes in order to reduce the dimensionality without reducing the classification accuracy. We first introduce the study context, before exposing preliminary studies on tuning the SVM learning method. The adapted method is then accurately exposed and the interesting experimental results as well as a sample of applications are presented before to conclude.
|
| [9] |
多源遥感影像决策级融合及其应用研究[D] .Multi-source remote sensing image fusion based on decision-level and its application[D]. |
| [10] |
Spatial location and ecological content of support vectors in an SVM classification of tropical vegetation [J].https://doi.org/10.1080/2150704X.2013.784848 Magsci 摘要
The Support Vector Machines (SVM) are increasingly far-reaching in remotely sensed data classification. As supervised classifiers, the SVM output depends on the input pixels, pointing out that training is potentially an important stage for optimizing classification accuracy. The SVM consist in projecting pixels into a high-dimensional feature space and then fitting in a hyperplane, maximizing the distance between the closest vectors and the hyperplane itself. This study aims to locate the pixels acting as support vectors and identify the ecological features they contain in a tropical vegetation context. The analyses focused on a Quickbird-2 image where two vegetation types occur. The physical boundary between classes was delineated in the field, and we used an iterative method to mark and localize the pixels acting as support vectors on the image. Our results highlight that vegetation sampling should focus on ecotones (the transition area between two different and adjacent vegetation classes) in order to minimize the field survey effort and maximize the mapping accuracy.
|
| [11] |
SVM spatio-temporal vegetation classification using HR satellite images [J].https://doi.org/10.1117/12.898256 URL [本文引用: 1] 摘要
This paper proposes a new HR spatio-temporal vegetation classification approach based on SVM. A multi-band SVM approach is first applied on satellite images time series then a graph based SVM algorithm is used for temporal analysis.
|
| [12] |
基于SVM决策支持树的城市植被类型遥感分类研究 [J].https://doi.org/10.11834/jrs.20060229 Magsci [本文引用: 1] 摘要
城市植被类型不同,生物量不同,其生态功能与绿化效应也不同。在目前难直接获取城市“绿量”实测数据的情况下,可以绿地面积和植被类型间接反映绿地的生物量和绿化效应。本文利用高分辨率卫星影像IKONOS,以实验区与验证区城市植被类型信息为对象,在对常用的参数和非参数分类方法进行对比实验的基础上,对SVM的核函数进行了分析,构建了基于SVM决策树的城市植被类型分类模型。分类实验结果表明:与其他传统方法分类结果比较,SVM的决策树分类方法对植被类型的分类精度达到83.5%,绿化面积总精度接近95%,取得了良好的效果。
Research on remote sensing classification of urban vegetation species based on SVM decision-making tree [J].https://doi.org/10.11834/jrs.20060229 Magsci [本文引用: 1] 摘要
城市植被类型不同,生物量不同,其生态功能与绿化效应也不同。在目前难直接获取城市“绿量”实测数据的情况下,可以绿地面积和植被类型间接反映绿地的生物量和绿化效应。本文利用高分辨率卫星影像IKONOS,以实验区与验证区城市植被类型信息为对象,在对常用的参数和非参数分类方法进行对比实验的基础上,对SVM的核函数进行了分析,构建了基于SVM决策树的城市植被类型分类模型。分类实验结果表明:与其他传统方法分类结果比较,SVM的决策树分类方法对植被类型的分类精度达到83.5%,绿化面积总精度接近95%,取得了良好的效果。
|
| [13] |
遥感图像森林林型SVM分类的多特征选择 [J].
为了研究遥感图像森林林型SVM分类多特征的选择对提高分类精度的影响,选取小波变换不同尺度纹理、四种植被指数、最优波段光谱特征等不同组合构成林型分类多特征向量进行分类。结果表明,纹理与植被指数、最优波段组合多特征的森林林型分类精度最高,阔叶林、针叶林和竹林的分类精度分别为84.4%、86.5%、91.0%,比纹理单类特征分类分别提高4.1%、4.0%、1.1%,比植被指数单类特征分类分别提高9.2%、11.8%、11.9%。多特征的分类精度一般要高于单类特征,纹理能够较明显提高林型可分性,植被指数也有一定的效果,但最优波段光谱特征的效果较弱。
Multi-feature selection in remote sensing forest species classification with SVM.
为了研究遥感图像森林林型SVM分类多特征的选择对提高分类精度的影响,选取小波变换不同尺度纹理、四种植被指数、最优波段光谱特征等不同组合构成林型分类多特征向量进行分类。结果表明,纹理与植被指数、最优波段组合多特征的森林林型分类精度最高,阔叶林、针叶林和竹林的分类精度分别为84.4%、86.5%、91.0%,比纹理单类特征分类分别提高4.1%、4.0%、1.1%,比植被指数单类特征分类分别提高9.2%、11.8%、11.9%。多特征的分类精度一般要高于单类特征,纹理能够较明显提高林型可分性,植被指数也有一定的效果,但最优波段光谱特征的效果较弱。
|
| [14] |
Although GIS provides powerful functionality for spatial analysis, data overlay and storage, these spatially oriented systems lack the ability to represent temporal dynamics, which is a major impediment to its use in surface modeling. However, rapid development of computing technology in recent years has made real-time spatial analysis and real-time data visualization become realizable. Based on newly developed methods, explores solutions to big-error and slow-efficiency problems, two critical challenges that have long plagued those working in with geographical information system (GIS) and computer-aided design (CAD).
|
| [15] |
Climate change trend in China: With improved accuracy [J].https://doi.org/10.1007/s10584-013-0785-5 Magsci [本文引用: 1] 摘要
We have found that a spatial interpolation of mean annual temperature (MAT) in China can be accomplished using a global ordinary least squares regression model since the relationship between temperature and its environmental determinants is constant. Therefore the estimation of MAT does not very across space and thus exhibits spatial stationarity. The interpolation of mean annual precipitation (MAP), however, is more complex and changes spatially as a function of topographic variation. Therefore, MAP shows spatial non-stationarity and must be estimated with a geographically weighted regression. A statistical transfer function (STF) of MAT was formulated using minimized residuals output from a high accuracy and high speed method for surface modeling (HASM) with an ordinary least squares (OLS) linear equation that uses latitude and elevation as independent variables, abbreviated as HASM-OLS. The STF of MAP under a BOX-COX transformation is derived as a combination of minimized residuals output by HASM with a geographically weighted regression (GWR) using latitude, longitude, elevation, impact coefficient of aspect and sky view factor as independent variables, abbreviated as HASM-GWR-BC. In terms of HASM-OLS and HASM-GWR-BC, MAT had an increasing trend since the 1960s in China, with an especially accelerated increasing trend since 1980. Overall, our data show that MAT has increased by 1.44 A degrees C since the 1960s. The warming rates increase from the south to north in China, except in the Qinghai-Xizang plateau. Specifically, the 2,100 A degrees C A center dot d contour line of annual accumulated temperature (AAT) of a parts per thousand yen10 A degrees C shifted northwestward 255 km in the Heilongjiang province since the 1960s. MAP in Qinghai-Xizang plateau and in arid region had a continuously increasing trend. In the other 7 regions of China, MAP shows both increasing and decreasing trends. On average, China became wetter from the 1960s to the 1990s, but drier from the 1990s to 2000s. The Qinghai-Xizang Plateau and Northern China experienced more climatic extremes than Southern China since the 1960s.
|

/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}