
Building Transfer Functions Between Testate amoeba and Environmental Variables with ‘rioja’ Package
Received date: 2013-01-02
Request revised date: 2013-04-17
Online published: 2013-08-20
Copyright
R language, as an open source programming language and software environment, is widely used in statistics for its free availability. The ‘rioja’ package of R specially deals with the analysis of Quaternary science data, containing functions for constrained clustering, transfer functions and plotting stratigraphic data. Testate amoebae are a group of unicellular protists living in terrestrial habitats. Their decayed resistant and morphologically diagnostic shells (tests) allow them to be extensive used as proxy in peat based paleoenvironmental reconstruction. This study aimed to: ① Present an example of application of ‘rioja’ package; ② build Testate amoeba-based transfer functions for quantitatively reconstructing paleoenvironment changes in Changbai Mountains with peat archive. The training set was constituted by 75 samples collected from four peatlands, Hani(42°12′50″N, 126°31′05″E), Jinchuan(42°20′47″N, 126°21′35″E), Chichi(42°03′16″N,128°03′22″E) and Yuanchi(42°01′55″N,128°25′58″E), in Changbai Mountains, northeast China. Three factors, depth to water table (DWT), pH and peat moisture, were selected as the target environmental variables. The models of Weighted Averaging (WA) and Weighted Averaging Partial Least Squares (WA-PLS) were used to build transfer functions. Leave-one-out was chosen as cross validation method. The results showed that the second component of WA-PLS is the best models for DWT producing a RMSEP of 7.39 and R2 of 0.74. For pH and peat moisture, both first component of WA-PLS and WA with inverse deshrinking could be regarded as the best models for they have the lowest RMSEP and relatively higher R2. The RMSEP of pH is 0.18 and R2 is 0.72, while for peat moisture RMSEP is1.95% and R2 is 0.62. The performances of the transfer function were comparable with other studies in the world. DWT, pH and peat moisture could be quantitive reconstructed with the mean errors of ±7.39 cm, ±0.18 and ±1.95%, respectively, if Testate amoebae assemblage of profiles was the same as the training set in this study.
Key words: ‘rioja’ package; transfer function; Testate amoebae; depth to water table; pH; peat moisture
LI Hong-kai , LI Wei-wei , PU You-bao , WANG Cong-yang , WANG Song-mei , YANG Xiao-lin . Building Transfer Functions Between Testate amoeba and Environmental Variables with ‘rioja’ Package[J]. SCIENTIA GEOGRAPHICA SINICA, 2013 , 33(8) : 1022 -1028 . DOI: 10.13249/j.cnki.sgs.2013.08.1022
Table 1 Predication performance of transfer functions for DWT, pH and peat moisture表1 水位埋深、pH和泥炭湿度转换函数预测性能 |
| 模型 | 水位埋深 | pH | 泥炭湿度 | |||
|---|---|---|---|---|---|---|
| RMSEP | R2 | RMSEP | R2 | RMSEP | R2 | |
| WA.inv | 10.93 (7.75) | 0.57 (0.72) | 0.29 (0.18) | 0.48 (0.72) | 3.40% (1.95%) | 0.34 (0.62) |
| WA.cla | 12.99 (8.31) | 0.59 (0.72) | 0.36 (0.19) | 0.50 (0.73) | 4.85% (2.26%) | 0.37 (0.63) |
| WA.inv.tol | 11.28 (8.33) | 0.55 (0.68) | 0.28 (0.18) | 0.51 (0.70) | 3.42% (1.97%) | 0.34 (0.62) |
| WA.cla.tol | 13.35 (9.10) | 0.56 (0.68) | 0.36 (0.23) | 0.52 (0.68) | 4.76% (2.19%) | 0.36 (0.62) |
| WA-PLS | 10.93 (7.39) | 0.57 (0.74) | 0.29 (0.18) | 0.48 (0.72) | 3.40% (1.95%) | 0.34 (0.62) |
Fig.1 Performance of transfer function models for DWT, pH and peat moisture, pre-( a,b,e,f,i,j) and post-( c,d,g,h,k,l) data filtering图1 优化前(a,b,e,f,i,j)后(c,d,g,h,k,l)水位埋深、pH和泥炭湿度转换函数模型的预测性能 |
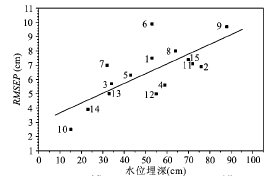
Fig.2 The RMSEP versus DWT gradient range in different studies图2 不同研究中的RMSEP与水位梯度宽度关系 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
R Core Team. R: A Language and Environment for Statistical Computing [CP/OL]. Vienna, Austria: R Foundation for Statistical Computing,2012[2012-12-15].
|
| [44] |
Juggins S. rioja: Analysis of Quaternary Science Data [CP/OL]. 2012 [2012-12-15].
|
| [45] |
|
| [46] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}