引言
包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世。防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] 。生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] 。研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素。其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] 。沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] 。本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制。
1 研究方法
1.1 研究区概况
包兰铁路沙坡头段防护体系位于腾格里沙漠东南缘(104°59′E,37°27′N),地势西北高东南低,沙丘类型以格状沙丘为主,同时广泛分布新月形沙丘和沙丘链。受蒙古高压和地形影响,该区主风向为NW和WNW,次主风向为ENE。气候干旱,年均降水量186.2 mm,年蒸发量高于3 000 mm。年均气温9.6 ℃,气温季节变化剧烈,7月平均气温24.3℃,1月平均气温- 6.9℃。年均风速3.5 m/s,春季大风频繁,风沙活动活跃。
铁路防护体系由砾石平台、灌溉植被带、无灌溉植被防护带和草方格沙障(1 m×1 m)、前沿阻沙栅栏组成。无灌溉植被防护带和草方格沙障是防护体系的主体部分。构成防护带的灌木主要有柠条(Caragana korshinskii, Leguminosae )、花棒(He
dysarum scoparium, Papilionaceae )、沙枣(angustifolia, Elaeagnaceae )、沙拐枣(Calligonum arborescens, Polygonaceae )。自1956年建立以来,防护带内不同发育程度的生物土壤结皮[9 ,22 ] ,提高了防护体系的防风固沙功能。
1.2 样品采集与测试
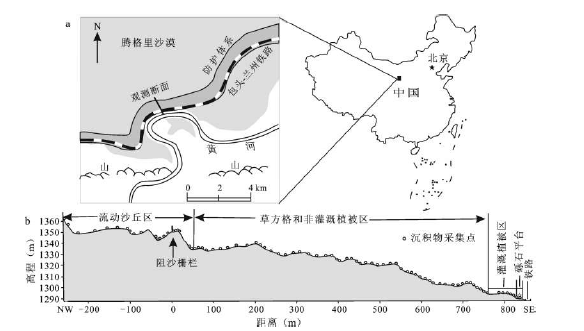
本文在防护体系内选取与主风向(NW)大致平行的断面。断面始于上风向防护体系外围、包含完整格状沙丘的流沙区,止于铁路以北的砾石平台(图1 ),全长约1 100 m。沿断面调查生物土壤结皮并采集地面沉积物样品。结皮调查包括结皮类型、厚度和紧实度。依据沙丘地貌部位变化,采用目测方法选择能够代表该沙丘地貌部位结皮分布状况的调查点和采样点,在各点布设1 m×1 m的“田”字样方,分别记录9个节点的结皮类型,测定结皮厚度和表层(0~5 cm)土壤紧实度,测定结果平均值分别作为该点结皮厚度和紧实度。根据该区域关于生物土壤结皮种类的研究[8 ] ,将结皮划分为苔藓结皮和地衣- 藻结皮。前者表面簇状分布具有茎叶的苔藓(图2 a),后者表面藻类和真菌形成的叶状体匍匐生长,形成凹凸的壳状覆盖(图 2 b)[23 ] 。结皮厚度使用精度为0.02 mm的游标卡尺测定。地表紧实度采用SC 900土壤紧实度仪(SC 900 Soil Compaction Meter,Spectrum Technologies Inc.,U.S.A.)测定。地表沉积物样点涵盖整个断面,其中防护体系外流沙区布设采样点16个,防护体系内流沙区(阻沙栅栏- 阻沙栅栏所在沙丘背风坡脚)布设采样点8个,流沙区下风向无生物土壤结皮分布的植被带布设采样点3个,有生物土壤结皮分布的区域布设采样点47个。流沙区采集0~5、5~10、10~15 cm共3层样品,其中沙丘背风坡仅采集表层样(0~5 cm);流沙区下风向各样点分结皮层、结皮层下至5、5~10、10~15、15~20 cm共5个采样深度,没有结皮分布的位置采集0~5、5~10、10~15、15~20 cm 4层样品。样品采集面积均为15 cm×15 cm,采样时首先目视选择样点周围1 m×1 m范围内代表性最好的结皮区域,依次采集各层样品。全部野外工作于2009年4~5月完成。
图1 研究区位置(a)与采样点(b)
Fig.1 Location of the study area (a) and the distribution of sampling sites (b)
图2 苔藓结皮(a)和地衣- 藻结皮(b)
Fig.2 The moss crust (a) and lichen-algae crust (b)
沉积物粒度采用Mastersizer 2000(Malvern Instruments,Malvern,UK)测定。粒度测试于2009年9~10月完成。本文采用Folk-Ward公式计算[24 ] 粒度参数,参照Blott和Pye[25 ] 使用的标准对粒度参数结果进行分级。
2 结果与分析
2.1 生物土壤结皮粒度参数及其分布
根据粒度测试结果,无论流沙区还是防护体系内部植被区,各采样点采样深度>5 cm各层沉积物的粒径组成、粒度参数以及各组分含量都很接近,故所有粒度参数均取5~20 cm深度范围内对应参数的平均值,与结皮层和结皮层下至5 cm范围内沉积物对应参数进行对比。
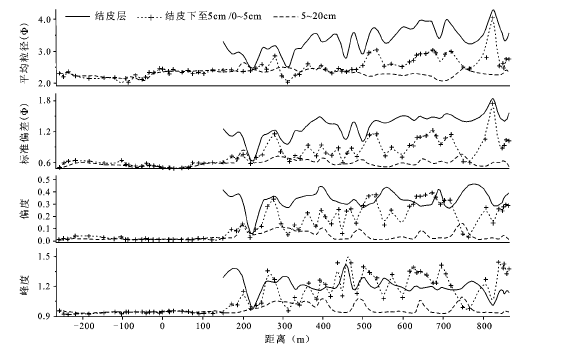
统计表明,防护体系内生物土壤结皮层颗粒平均粒径为3.59 Ф,结皮层下至5 cm平均粒径为2.63 Ф,5~20 cm为2.36 Ф;流沙区表层(0~5 cm)沉积颗粒平均粒径为2.28 Ф,5~20 cm为2.25 Ф。可见植被带内有结皮分布的区域,结皮层沉积物颗粒比下层普遍变细(图3 )。结皮层沉积颗粒总体分选差,极细偏(0.33),极单峰尖窄;结皮层下沉积物颗粒中等分选,细偏(0.09),单峰尖窄;流沙区颗粒分选中等偏好,正态分布(0.02),常峰态。因此防护体系结皮层沉积物粒度参数与结皮层下沉积物、流沙区沉积物相比差异显著,结皮层下沉积物和流沙区沉积物粒度参数差异不显著,但前者平均粒径略细,分选稍差。
水平方向上,流沙区各层沉积物粒径随距离变化小,结皮层粒径变化显著,结皮层下至5 cm沉积物粒径变化较明显。5~20 cm深度沉积物在整个断面上变化微弱。相同采样深度内,防护体系内沉积物比流沙区细。随防护距离增大,防护体系内生物土壤结皮层和结皮层下至5 cm沉积物粒径表现出波动性变细特征,结皮层的波动性尤为明显。防护体系内固定或半固定沙丘丘顶位置表层沉积物颗粒相对较粗,分选性较好;丘间地表层沉积物颗粒较细,分选性差。流沙区沉积物和防护体系内的下层沉积物粒径随地形变化未表现出明显的波动性变化特征(图3 )。
图3 沉积物粒度参数沿断面变化
Fig.3 Variations of grain-size parameters along the cross-section
2.2 生物土壤结皮组分含量变化
整个断面上粗砂含量低(<0.4%),粘粒含量较少(<5.2%)。细砂是沉积物的主要组分,防护体系内,细砂含量从结皮层的59.1%增加到结皮层下至5 cm深度的67.2%,5~20 cm深度达到70.0%;流沙区细砂含量从表层(0~5 cm)的70.2%降低到下层(5~20 cm)的68.4%。防护体系内结皮层粘粒含量(1.5%)高于结皮下至5 cm(0.3%),深度>5 cm不含粘粒。结皮层粉砂含量(27.3%)则远高于结皮下至5cm(9.3%)和5~20 cm(1.4%);流沙区表层及其下层不含粘粒和粉砂。防护体系内中砂含量从结皮层的12.1%增加到结皮下至5 cm的23.5%,5~20 cm深度增至27.4%;流沙区中砂含量从表层的29.4%增加到下层的31.5%。流沙区细砂和中砂含量略高于防护体系,含微量粗砂,未检测到粘粒和粉砂。防护体系内粘粒和粉砂是结皮层沉积物的重要组分。无论流沙区还是植被带,沉积物中各组分含量均表现出随深度增大,细组分含量降低,而粗组分含量增加的趋势。随防护距离增大,各组分含量均表现出随地形变化的波动性。其中,细组分(粉砂和粘粒)丘顶最少,丘间地增多,随防护距离增大而波动性增大;粗组分(细砂和中砂)呈现相反的变化趋势。流沙区细砂和中砂含量也表现出相反的波动变化特征(图4 )。
灌溉植被区沉积物平均粒径最细,分选最差,细偏程度大,颗粒粒径分布集中程度较小(峰度为常峰态)。沉积物粒度分布异常的一个重要原因是该区域表土有客土成分,非风沙活动中的自然沉积。
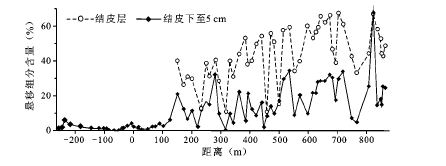
防护体系内结皮层是风沙活动减弱后逐渐形成的,其粒度组成变化直接反映了风沙活动方式的改变,因此对局地风沙环境变迁具有重要的指示意义。本文将表层(0~5 cm)沉积物划分为悬移(<0.1 mm)、跃移(0.1~0.5 mm)和蠕移颗粒(0.5~1.0 mm)[26 ] ,依据表层沉积物中悬移、跃移和蠕移组分的含量变化,推溯防护体系内风沙活动方式的转变规律。结果显示,跃移颗粒仍是防护体系表层沉积物的主体组分(75.2%),悬移颗粒随防护距离增大而逐渐成为重要组分(24.7%)(图5 ),蠕移颗粒极有限。结皮层中的悬移和跃移颗粒含量及其空间变化都与结皮层下至5 cm层以及流沙区存在明显差异。结皮层悬移含量略高于跃移颗粒,结皮层下至5 cm层跃移颗粒是主体(82.4%)。流沙区表层跃移颗粒占绝对优势(97.8%)。空间分布上,随防护距离增大,结皮层中悬移颗粒含量波动性增加,跃移颗粒含量波动性减小。流沙区沉积物和防护体系内结皮层下至5 cm层中,跃移和悬移颗粒含量变化都很小。灌溉植被区由于地表人为客土,表层沉积物中悬移颗粒含量极高。
图4 沉积物组分含量沿断面变化
Fig.4 Variations of sediment composition along the cross-section
图5 结皮层和结皮层下至5 cm沉积物中悬移组分含量
Fig.5 Content of suspended components in the sediment of biological soil crust layer and the content in 5 cm under crust layer
2.3 生物土壤结皮类型和地表紧实度变化
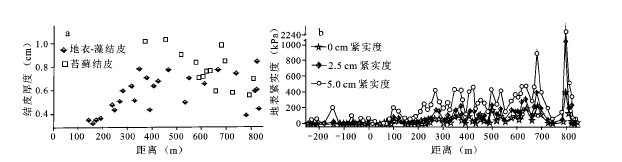
防护体系内防护距离<150 m的区域未发育生物土壤结皮。地衣- 藻结皮和苔藓结皮分布范围和厚度都明显不同。地衣- 藻结皮广泛分布,平均厚度0.576 cm;苔藓结皮主要分布在防护距离较大的区域,平均厚度0.796 cm,显著大于地衣- 藻结皮厚度。防护距离150~270 m范围内,地衣- 藻结皮呈斑块状分布,270 m以远的区域除部分固定沙丘丘顶无分布或斑块状分布外,其余区域基本为连续分布。地衣- 藻结皮厚度随防护距离增加逐渐变厚,苔藓结皮厚度无明显变化趋势。在靠近铁路的区域,由于人为干扰频繁,结皮发育受到影响,厚度较小(图6 a)。在苔藓结皮分布区域,苔藓结皮主要分布在丘间地和平缓坡脚区域,地衣- 藻结皮主要分布在沙丘的中上部和低矮固定沙丘丘顶。
由于结皮层大多很薄,直接测定其紧实度非常困难,本文以表层(0~5 cm)紧实度反映结皮紧实度。结果表明,流沙区表层紧实度很小,平均仅为10.9 kPa;防护体系内除灌溉林地紧实度异常大外,其他区域结皮紧实度介于3.9~896.9 kPa。随防护距离增大,结皮紧实度表现出一定的增大趋势(图6 b)。
图6 结皮类型、厚度(a)与地表紧实度(b)沿断面的分布
Fig.6 Distribution of crust type, thickness (a), and ground compaction (b) along the cross-section
3 讨 论
.1 生物土壤结皮粒度变化环境意义
防护体系内表层尤其是生物土壤结皮层沉积物较流沙区细,分选性差。随防护距离增大,表层(结皮层和结皮层下至5 cm层)沉积物粒径逐渐变细,同时表现出波动性变化。深度>5 cm沉积物粒度在整个断面上变化微弱,表明深度5 cm以下的沉积物主要是防护体系建立前剧烈风沙活动的结果。结皮层中粘粒和粉砂含量远高于其它层,两者含量随深度增加急剧减少。流沙区沉积物主要由细砂和中砂组成,细砂是主要组分,各层均未检测到粘粒和粉砂,表明防护体系内的粘粒和粉砂并非来源于上风向邻近的流沙区,而可能来源于远距离悬移沉降物。粘粒、粉砂的积累和结皮层的发育,标志着防护体系风沙环境性质发生了根本改变。植被带前段区域结皮发育,颗粒运动方式以跃移为主,但表层沉积物中悬移组分含量已远大于流沙区,表明防护体系前段地表风蚀、搬运过程逐渐减弱,上风向风沙流携带的粗组分迅速沉积,较细组分被搬运至较远距离沉积。该区域下风向植被带风蚀微弱,风沙活动方式转变为以大气降尘和近地面悬移颗粒的绝对沉积为主,为结皮的发育营造了稳定的地表条件。
3.2 生物土壤结皮形成发育和地表紧实度环境意义
防护体系前段是风沙活动由强烈风蚀和堆积向沉积为主转变的过渡区域,地表的稳定性不能满足结皮发育,因而没有结皮分布。研究表明地衣-藻结皮是生物土壤结皮的先驱类型,其先于苔藓结皮发育,苔藓结皮是生物土壤结皮发育的成熟阶段[27 ~29 ] 。地衣喜少干扰、相对稳定的土壤,其发育还受景观稳定性制约,可作为生态环境变化的指示[6 ] 。苔藓结皮仅分布在防护体系防护距离较大,植被带建立较早的区域,表明该区域地表稳定性很好,非常利于结皮发育成熟。地衣- 藻结皮在防护体系内广泛分布,表明这些区域地表处于相对稳定状态,为结皮发育提供了最基本的地表稳定条件。苔藓结皮发育区域,苔藓结皮和地衣- 藻结皮分布在固定沙丘的不同地貌部位,这是生物对环境条件适应的一种生态表现[30 ] 。研究证实生物土壤结皮可以显著提高地表的抗风蚀能力,苔藓结皮的抗风蚀能力明显高于地衣- 藻结皮[31 ] ,综上所述,两种生物土壤结皮在防护体系内的差异性分布特征指示了防护体系内地表抗风蚀能力和风沙环境稳定性的空间差异,即沿断面地表稳定性逐渐增强,地势较低和平缓的沙丘地貌部位地表相对更为稳定。
紧实度可以衡量地表抵抗外力的能力,紧实度越大,地表越不易遭受外力破坏。防护体系内表层紧实度远大于流沙区,表明结皮层对增强地表的抗风蚀能力发挥着重要作用。但防护体系内地表紧实度总体仍处于较低水平,表明结皮层仍比较脆弱,容易破损。随防护距离增大,结皮发育程度逐渐变好,地表紧实度逐渐增大,指示了随防护距离增大,地表抗风蚀能力逐渐增强,稳定性逐渐变好。紧实度随地形起伏而表现出的波动性再次表明结皮的发育受地形变化的制约,在不同沙丘部位,结皮的抗风蚀能力存在差异。丘顶结皮紧实度较小,抗风蚀能力较弱,丘间地结皮紧实度较大,抗风蚀能力相对较强。
防护体系内表层沉积物的粒度变化和生物土壤结皮的空间差异表明,沙区风沙环境变化主导着土壤形成发育的方向和速度。防护体系(特别是其中的人工植被和草方格)营造沉积主导的风沙环境,为远距离输送的悬移细颗粒的沉降和富集提供了有利条件,后者则为流沙表面生物土壤结皮的形成发育提供了极其重要的物质基础。粘粒含量是影响土壤肥力的主要因素之一[32 ] ,结皮层中粘粒含量明显提高,既证实沙尘沉降与富集主导的风沙环境的形成,也预示土壤肥力的逐步提高和土壤发育环境的改善。本文结果显示,苔藓结皮和地衣- 藻结皮在沙丘不同地貌部位的分布格局,以及结皮层粘粒含量的变化特征,都与王雪芹等[30 ] 在古尔班通古特沙漠和段争虎等[33 ] 在腾格里沙漠的相关研究结果类似,可见具有普遍意义的是,流沙地区风沙环境性质的改变及其程度和沙丘地形的起伏,是生物土壤结皮形成发育和空间分异的根本原因,而结皮沉积物粒度特征的水平差异性和剖面变化则忠实记录和指示风沙环境的时空变化。
4 结 论
沙坡头铁路防护体系内,由西北向东南沿主风向从无结皮分布逐渐变为结皮斑块状分布,再发展到结皮连续分布;结皮类型由地衣- 藻结皮为主过渡到苔藓结皮和地衣- 藻结皮交错分布,地表紧实度随防护距离呈增大趋势。表层尤其是结皮层沉积物粒径逐渐变细,分选变差。细砂仍是防护体系内沉积物的主要组分,但表层粘粒和粉砂含量既高于流沙区也高于防护体系内表层以下各层,且表层沉积物中细组分(粘粒和粉砂)含量随防护距离增大而增多,粗组分(细沙和中砂)含量相对减少。流沙区沉积物中跃移颗粒占绝对优势,防护体系内即使是无结皮分布的区域,悬移颗粒含量也远高于流沙区,且随防护距离的增大表现出增加趋势。受防护体系内固定和半固定沙丘地形影响,结皮厚度、类型、地表紧实度、表层沉积物的机械组成、跃移和悬移组分含量变化还呈现出一定的波动性。
沉积物粒度组成和结皮的分布规律表明随防护距离的增大地表稳定性逐渐增强,风力搬运能力逐渐减弱。流沙区普遍存在强烈的蚀积过程,防护体系前段150 m范围内风蚀迅速削弱,跃移颗粒的沉积成为主导风沙过程,150 m以远区域发展为绝对沉积区,沉积颗粒以大气降尘和近地面悬移颗粒为主。风沙活动方式的转变表明在防护体系内风沙环境趋于稳定。
致 谢:中国科学院寒区旱区环境与工程研究所沙坡头沙漠试验研究站李新荣研究员、樊恒文高级工程师对本项工作提供了热情帮助和支持,参加野外调查采样的还有刘永刚、周娜,粒度测试实验得到马晓洁和渠敏的帮助,特致谢忱!
The authors have declared that no competing interests exist.
参考文献
文献选项
[1]
甄计国 . 腾格里沙漠东南缘沙坡头地区流沙治理后地表形态的变化
[J].中国沙漠 ,1987 ,7 (1 ):9 ~17 .
[本文引用: 1]
[2]
张春来 ,邹学勇 ,程宏 ,等 .包兰铁路沙坡头段防护体系近地面流场特征
[J].应用基础与工程科学学报 ,2006 ,14 (3 ):353 ~360 .
[本文引用: 1]
[3]
屈建军 ,凌裕泉 ,井哲帆 ,等 .包兰铁路沙坡头段风沙运动规律及其与防护体系的相互作用
[J].中国沙漠 ,2007 ,27 (4 ):529 ~533 .
[本文引用: 1]
[4]
刘媖心 . 包兰铁路沙坡头地段铁路防沙体系的建立及其效益
[J].中国沙漠 ,1987 ,7 (4 ):1 ~11 .
[本文引用: 1]
[5]
段争虎 ,刘新民 ,屈建军 .沙坡头地区土壤结皮形成机理的研究
[J].干旱区研究 ,1996 ,13 (2 ):31 ~36 .
[本文引用: 1]
[6]
李新荣 ,贾玉奎 ,龙利群 ,等 .干旱半干旱地区土壤微生物结皮的生态学意义及若干研究进展
[J].中国沙漠 ,2001 ,21 (1 ):4 ~11 .
[本文引用: 1]
[7]
李守中 ,郑怀周 ,李守丽 ,等 .沙坡头植被固沙区生物结皮的发育特征
[J].生态学杂志 ,2008 ,27 (10 ):1675 ~1679 .
[8]
邵玉琴 ,赵吉 ,包青海 ,等 .沙坡头固定沙丘结皮层的微生物区系动态
[J].中国沙漠 ,2002 ,22 (3 ):298 ~303 .
[本文引用: 2]
[9]
张继贤 ,邸醒民 ,王淑湘 .沙坡头地区防护体系建立过程中生态环境变化的特点
[J].干旱区资源与环境 ,1994 ,8 (3 ):68 ~79 .
[本文引用: 2]
[10]
Tsoar H Pye K. Dust transport and the question of desert loess formation
[J].Sedimentol ,1987 ,34 (1 ):139 -153 .
[本文引用: 1]
[11]
Pye K. Aeolian dust and dust deposits [M].London :Academic Press ,1987 :334 .
[本文引用: 1]
[12]
徐佳 ,刘普灵 ,邓瑞芬 ,等 .黄土坡面不同植被恢复阶段的减流减沙效益研究
[J].地理科学 ,2012 ,32 (11 ):1391 ~1396 .
[本文引用: 1]
[13]
Rosenberg N J. Microclimate: the biological environment
[M].New York:Wiley ,1974 .
[本文引用: 1]
[14]
何兴东 ,赵爱国 ,段争虎 ,等 .塔里木沙漠公路灌木固沙带的防护效应
[J].生态学杂志 ,2002 ,21 (4 ):26 ~30 .
[15]
汪万福 ,王涛 ,李最雄 ,等 .敦煌莫高窟崖顶灌木林带防风固沙效应
[J].生态学报 ,2004 ,24 (11 ):2492 ~2500 .
[本文引用: 1]
[16]
舒强 ,陈晔 ,张茂恒 ,等 .苏北平原周庄镇浅钻沉积物粘土矿物分布特征及其环境意义
[J].地理科学 ,2012 ,32 (2 ):238 ~243 .
[本文引用: 1]
[17]
Visher G S. Grain size distributions and depositional processes
[J].J. Sediment. Petrol. ,1969 ,39 (3 ):1074 -1106 .
[18]
何清 ,杨兴华 ,霍文 ,等 .库姆塔格沙漠粒度分布特征及环境意义
[J].中国沙漠 ,2009 ,29 (1 ):9 ~13 .
[本文引用: 1]
[19]
Pye K Tsoar H. Aeolian sand and sand dunes
[M].London:Unwin Hyman ,1990 .
[本文引用: 1]
[20]
Winspear N R Pye K. Sand supply to the Algodones dunefield, sou-
theastern California USA [J].Sedimentol. ,1995 ,42 (6 ):875 -891 .
[21]
周娜 ,张春来 ,刘永刚 .雅鲁藏布江米林宽谷段新月形沙丘粒度分异研究
[J].地理科学 ,2011 ,31 (8 ):958 ~963 .
[本文引用: 1]
[22]
刘家琼 . 草原与荒漠交界地区流沙的固定——以沙坡头铁路防护体系旱路固沙为例
[J].中国沙漠 ,2002 ,22 (5 ):498 ~498 .
[本文引用: 1]
[23]
王雪芹 ,张元明 ,张伟民 ,等 .生物结皮粗糙度特征——以古尔班通古特沙漠为例
[J].生态学报 ,2011 ,31 (14 ):4153 ~4160 .
[本文引用: 1]
[24]
Folk R L Ward W C. Brazos River bar, a study in the significance of grain-size parameters
[J].J. Sediment. Petrol. ,1957 ,27 (1 ):3 -26 .
[本文引用: 1]
[25]
Blott S J Pye K. Gradistat:A grain size distribution and statistics package for the analysis of unconsolidated sediments
[J].Earth Surf. Process. Landforms ,2001 ,26 (11 ):1237 -1248 .
[本文引用: 1]
[26]
Hansen Z K Libecap G D. Small farms, externalities, and the dust bowl of the 1930s
[J].J. Polit. Econ. ,2004 ,112 (3 ):665 -694 .
[本文引用: 1]
[27]
Johansen J R. Minireview: cryptogamic crusts of semiarid and arid lands of North America
[J].J. Psychol. , 1993 ,29 (2 ):140 -147 .
[本文引用: 1]
[28]
West N E. Structure and function of microphytic soil crusts in wildland ecosystems of arid to semi-arid regions
[J].Adv. Ecol. Res. ,1990 ,20 :179 -223 .
[29]
Yeager C M Kornosky J L Housman D C et al. Diazotrophic community structure and function in two successional stages of biological soil crusts from the Colorado Plateau and Chihuahuan Desert
[J].App. Environ. Microbiol. ,2004 ,70 (2 ):973 -983 .
[本文引用: 1]
[30]
王雪芹 ,张元明 ,王远超 ,等 .古尔班通古特沙漠生物结皮小尺度分异的环境特性
[J].中国沙漠 ,2006 ,26 (5 ):711 ~716 .
[本文引用: 2]
[31]
王雪芹 ,张元明 ,张伟民 ,等 .古尔班通古特沙漠生物结皮对地表风蚀作用影响的风洞实验
[J].冰川冻土 ,2004 ,26 (5 ):632 ~638 .
[本文引用: 1]
[32]
张丽萍 ,王小云 ,张赫斯 .沙盖黄土丘陵坡地土壤理化特性随地形变化规律研究
[J].地理科学 ,2011 ,32 (2 ):178 ~183 .
[本文引用: 1]
[33]
Duan Z H Xiao H L Li X R et al. Evolution of soil properties on stabilized sands in the Tengger Desert, China
[J].Geomorphol. ,2004 ,59 :237 -246 .
[本文引用: 1]
腾格里沙漠东南缘沙坡头地区流沙治理后地表形态的变化
1
1987
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
包兰铁路沙坡头段防护体系近地面流场特征
1
2006
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
包兰铁路沙坡头段风沙运动规律及其与防护体系的相互作用
1
2007
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
包兰铁路沙坡头地段铁路防沙体系的建立及其效益
1
1987
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
沙坡头地区土壤结皮形成机理的研究
1
1996
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
干旱半干旱地区土壤微生物结皮的生态学意义及若干研究进展
1
2001
... 防护体系前段是风沙活动由强烈风蚀和堆积向沉积为主转变的过渡区域,地表的稳定性不能满足结皮发育,因而没有结皮分布.研究表明地衣-藻结皮是生物土壤结皮的先驱类型,其先于苔藓结皮发育,苔藓结皮是生物土壤结皮发育的成熟阶段[27 ~29 ] .地衣喜少干扰、相对稳定的土壤,其发育还受景观稳定性制约,可作为生态环境变化的指示[6 ] .苔藓结皮仅分布在防护体系防护距离较大,植被带建立较早的区域,表明该区域地表稳定性很好,非常利于结皮发育成熟.地衣- 藻结皮在防护体系内广泛分布,表明这些区域地表处于相对稳定状态,为结皮发育提供了最基本的地表稳定条件.苔藓结皮发育区域,苔藓结皮和地衣- 藻结皮分布在固定沙丘的不同地貌部位,这是生物对环境条件适应的一种生态表现[30 ] .研究证实生物土壤结皮可以显著提高地表的抗风蚀能力,苔藓结皮的抗风蚀能力明显高于地衣- 藻结皮[31 ] ,综上所述,两种生物土壤结皮在防护体系内的差异性分布特征指示了防护体系内地表抗风蚀能力和风沙环境稳定性的空间差异,即沿断面地表稳定性逐渐增强,地势较低和平缓的沙丘地貌部位地表相对更为稳定. ...
沙坡头固定沙丘结皮层的微生物区系动态
2
2002
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
... 本文在防护体系内选取与主风向(NW)大致平行的断面.断面始于上风向防护体系外围、包含完整格状沙丘的流沙区,止于铁路以北的砾石平台(图1 ),全长约1 100 m.沿断面调查生物土壤结皮并采集地面沉积物样品.结皮调查包括结皮类型、厚度和紧实度.依据沙丘地貌部位变化,采用目测方法选择能够代表该沙丘地貌部位结皮分布状况的调查点和采样点,在各点布设1 m×1 m的“田”字样方,分别记录9个节点的结皮类型,测定结皮厚度和表层(0~5 cm)土壤紧实度,测定结果平均值分别作为该点结皮厚度和紧实度.根据该区域关于生物土壤结皮种类的研究[8 ] ,将结皮划分为苔藓结皮和地衣- 藻结皮.前者表面簇状分布具有茎叶的苔藓(图2 a),后者表面藻类和真菌形成的叶状体匍匐生长,形成凹凸的壳状覆盖(图 2 b)[23 ] .结皮厚度使用精度为0.02 mm的游标卡尺测定.地表紧实度采用SC 900土壤紧实度仪(SC 900 Soil Compaction Meter,Spectrum Technologies Inc.,U.S.A.)测定.地表沉积物样点涵盖整个断面,其中防护体系外流沙区布设采样点16个,防护体系内流沙区(阻沙栅栏- 阻沙栅栏所在沙丘背风坡脚)布设采样点8个,流沙区下风向无生物土壤结皮分布的植被带布设采样点3个,有生物土壤结皮分布的区域布设采样点47个.流沙区采集0~5、5~10、10~15 cm共3层样品,其中沙丘背风坡仅采集表层样(0~5 cm);流沙区下风向各样点分结皮层、结皮层下至5、5~10、10~15、15~20 cm共5个采样深度,没有结皮分布的位置采集0~5、5~10、10~15、15~20 cm 4层样品.样品采集面积均为15 cm×15 cm,采样时首先目视选择样点周围1 m×1 m范围内代表性最好的结皮区域,依次采集各层样品.全部野外工作于2009年4~5月完成. ...
沙坡头地区防护体系建立过程中生态环境变化的特点
2
1994
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
... dysarum scoparium, Papilionaceae )、沙枣(angustifolia, Elaeagnaceae )、沙拐枣(Calligonum arborescens, Polygonaceae ).自1956年建立以来,防护带内不同发育程度的生物土壤结皮[9 ,22 ] ,提高了防护体系的防风固沙功能. ...
Dust transport and the question of desert loess formation
1
1987
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
1
1987
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
黄土坡面不同植被恢复阶段的减流减沙效益研究
1
2012
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
Microclimate: the biological environment
1
1974
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
敦煌莫高窟崖顶灌木林带防风固沙效应
1
2004
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
苏北平原周庄镇浅钻沉积物粘土矿物分布特征及其环境意义
1
2012
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
Grain size distributions and depositional processes
0
1969
库姆塔格沙漠粒度分布特征及环境意义
1
2009
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
Aeolian sand and sand dunes
1
1990
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
雅鲁藏布江米林宽谷段新月形沙丘粒度分异研究
1
2011
... 包兰铁路沙坡头段防护体系以其卓越的风沙防护性能闻名于世.防护体系建成后地表形态[1 ] 、近地面流场[2 ] 、地表沉积、风沙活动形式与强度[3 ] 都发生显著改变,流动沙丘转变成为固定和半固定沙丘[4 ] .生物土壤结皮是防护体系建立后逐渐形成发育的,是防护体系效益的显著标志之一,众多学者围绕其发育机理和过程、微生物种类和数量、生态环境功能等方面开展大量研究[5 ~8 ] ,但生物土壤结皮抗风蚀能力及其与风沙环境变迁的相互作用研究成果还很不足[9 ] .研究表明,地形和植被状况直接影响地表稳定性[10 ] ,而后者是决定防护体系内土壤生物和沉积物分布的主要因素.其中地形主要通过影响风速和流场作用于沉积物分布[11 ] ,植被则通过增大地表粗糙度削弱近地表风速,制约风沙活动强度,从而增强地表稳定性和抗侵蚀能力[12 ] ,促进生物土壤结皮的发育[13 ~15 ] .沉积物特性对环境具有重要的指示意义,其中沉积物粒度分布的指示作用应用最为广泛[16 ~18 ] ,沙丘表面沉积物粒度分布是研究沙丘地貌及其发育的动力过程、回溯沙丘发育历史、区分风沙环境类型的重要依据[19 ~21 ] .本文拟通过分析防护体系内外生物土壤结皮类型、厚度,地表沉积物粒度组成和地表紧实度的空间差异,探讨防护体系内风沙环境稳定性的空间变化规律与成因,以及生物土壤结皮形成发育与风沙环境变迁的相互影响机制. ...
草原与荒漠交界地区流沙的固定——以沙坡头铁路防护体系旱路固沙为例
1
2002
... dysarum scoparium, Papilionaceae )、沙枣(angustifolia, Elaeagnaceae )、沙拐枣(Calligonum arborescens, Polygonaceae ).自1956年建立以来,防护带内不同发育程度的生物土壤结皮[9 ,22 ] ,提高了防护体系的防风固沙功能. ...
生物结皮粗糙度特征——以古尔班通古特沙漠为例
1
2011
... 本文在防护体系内选取与主风向(NW)大致平行的断面.断面始于上风向防护体系外围、包含完整格状沙丘的流沙区,止于铁路以北的砾石平台(图1 ),全长约1 100 m.沿断面调查生物土壤结皮并采集地面沉积物样品.结皮调查包括结皮类型、厚度和紧实度.依据沙丘地貌部位变化,采用目测方法选择能够代表该沙丘地貌部位结皮分布状况的调查点和采样点,在各点布设1 m×1 m的“田”字样方,分别记录9个节点的结皮类型,测定结皮厚度和表层(0~5 cm)土壤紧实度,测定结果平均值分别作为该点结皮厚度和紧实度.根据该区域关于生物土壤结皮种类的研究[8 ] ,将结皮划分为苔藓结皮和地衣- 藻结皮.前者表面簇状分布具有茎叶的苔藓(图2 a),后者表面藻类和真菌形成的叶状体匍匐生长,形成凹凸的壳状覆盖(图 2 b)[23 ] .结皮厚度使用精度为0.02 mm的游标卡尺测定.地表紧实度采用SC 900土壤紧实度仪(SC 900 Soil Compaction Meter,Spectrum Technologies Inc.,U.S.A.)测定.地表沉积物样点涵盖整个断面,其中防护体系外流沙区布设采样点16个,防护体系内流沙区(阻沙栅栏- 阻沙栅栏所在沙丘背风坡脚)布设采样点8个,流沙区下风向无生物土壤结皮分布的植被带布设采样点3个,有生物土壤结皮分布的区域布设采样点47个.流沙区采集0~5、5~10、10~15 cm共3层样品,其中沙丘背风坡仅采集表层样(0~5 cm);流沙区下风向各样点分结皮层、结皮层下至5、5~10、10~15、15~20 cm共5个采样深度,没有结皮分布的位置采集0~5、5~10、10~15、15~20 cm 4层样品.样品采集面积均为15 cm×15 cm,采样时首先目视选择样点周围1 m×1 m范围内代表性最好的结皮区域,依次采集各层样品.全部野外工作于2009年4~5月完成. ...
Brazos River bar, a study in the significance of grain-size parameters
1
1957
... 沉积物粒度采用Mastersizer 2000(Malvern Instruments,Malvern,UK)测定.粒度测试于2009年9~10月完成.本文采用Folk-Ward公式计算[24 ] 粒度参数,参照Blott和Pye[25 ] 使用的标准对粒度参数结果进行分级. ...
Gradistat:A grain size distribution and statistics package for the analysis of unconsolidated sediments
1
2001
... 沉积物粒度采用Mastersizer 2000(Malvern Instruments,Malvern,UK)测定.粒度测试于2009年9~10月完成.本文采用Folk-Ward公式计算[24 ] 粒度参数,参照Blott和Pye[25 ] 使用的标准对粒度参数结果进行分级. ...
Small farms, externalities, and the dust bowl of the 1930s
1
2004
... 防护体系内结皮层是风沙活动减弱后逐渐形成的,其粒度组成变化直接反映了风沙活动方式的改变,因此对局地风沙环境变迁具有重要的指示意义.本文将表层(0~5 cm)沉积物划分为悬移(<0.1 mm)、跃移(0.1~0.5 mm)和蠕移颗粒(0.5~1.0 mm)[26 ] ,依据表层沉积物中悬移、跃移和蠕移组分的含量变化,推溯防护体系内风沙活动方式的转变规律.结果显示,跃移颗粒仍是防护体系表层沉积物的主体组分(75.2%),悬移颗粒随防护距离增大而逐渐成为重要组分(24.7%)(图5 ),蠕移颗粒极有限.结皮层中的悬移和跃移颗粒含量及其空间变化都与结皮层下至5 cm层以及流沙区存在明显差异.结皮层悬移含量略高于跃移颗粒,结皮层下至5 cm层跃移颗粒是主体(82.4%).流沙区表层跃移颗粒占绝对优势(97.8%).空间分布上,随防护距离增大,结皮层中悬移颗粒含量波动性增加,跃移颗粒含量波动性减小.流沙区沉积物和防护体系内结皮层下至5 cm层中,跃移和悬移颗粒含量变化都很小.灌溉植被区由于地表人为客土,表层沉积物中悬移颗粒含量极高. ...
Minireview: cryptogamic crusts of semiarid and arid lands of North America
1
1993
... 防护体系前段是风沙活动由强烈风蚀和堆积向沉积为主转变的过渡区域,地表的稳定性不能满足结皮发育,因而没有结皮分布.研究表明地衣-藻结皮是生物土壤结皮的先驱类型,其先于苔藓结皮发育,苔藓结皮是生物土壤结皮发育的成熟阶段[27 ~29 ] .地衣喜少干扰、相对稳定的土壤,其发育还受景观稳定性制约,可作为生态环境变化的指示[6 ] .苔藓结皮仅分布在防护体系防护距离较大,植被带建立较早的区域,表明该区域地表稳定性很好,非常利于结皮发育成熟.地衣- 藻结皮在防护体系内广泛分布,表明这些区域地表处于相对稳定状态,为结皮发育提供了最基本的地表稳定条件.苔藓结皮发育区域,苔藓结皮和地衣- 藻结皮分布在固定沙丘的不同地貌部位,这是生物对环境条件适应的一种生态表现[30 ] .研究证实生物土壤结皮可以显著提高地表的抗风蚀能力,苔藓结皮的抗风蚀能力明显高于地衣- 藻结皮[31 ] ,综上所述,两种生物土壤结皮在防护体系内的差异性分布特征指示了防护体系内地表抗风蚀能力和风沙环境稳定性的空间差异,即沿断面地表稳定性逐渐增强,地势较低和平缓的沙丘地貌部位地表相对更为稳定. ...
Structure and function of microphytic soil crusts in wildland ecosystems of arid to semi-arid regions
0
1990
Diazotrophic community structure and function in two successional stages of biological soil crusts from the Colorado Plateau and Chihuahuan Desert
1
2004
... 防护体系前段是风沙活动由强烈风蚀和堆积向沉积为主转变的过渡区域,地表的稳定性不能满足结皮发育,因而没有结皮分布.研究表明地衣-藻结皮是生物土壤结皮的先驱类型,其先于苔藓结皮发育,苔藓结皮是生物土壤结皮发育的成熟阶段[27 ~29 ] .地衣喜少干扰、相对稳定的土壤,其发育还受景观稳定性制约,可作为生态环境变化的指示[6 ] .苔藓结皮仅分布在防护体系防护距离较大,植被带建立较早的区域,表明该区域地表稳定性很好,非常利于结皮发育成熟.地衣- 藻结皮在防护体系内广泛分布,表明这些区域地表处于相对稳定状态,为结皮发育提供了最基本的地表稳定条件.苔藓结皮发育区域,苔藓结皮和地衣- 藻结皮分布在固定沙丘的不同地貌部位,这是生物对环境条件适应的一种生态表现[30 ] .研究证实生物土壤结皮可以显著提高地表的抗风蚀能力,苔藓结皮的抗风蚀能力明显高于地衣- 藻结皮[31 ] ,综上所述,两种生物土壤结皮在防护体系内的差异性分布特征指示了防护体系内地表抗风蚀能力和风沙环境稳定性的空间差异,即沿断面地表稳定性逐渐增强,地势较低和平缓的沙丘地貌部位地表相对更为稳定. ...
古尔班通古特沙漠生物结皮小尺度分异的环境特性
2
2006
... 防护体系前段是风沙活动由强烈风蚀和堆积向沉积为主转变的过渡区域,地表的稳定性不能满足结皮发育,因而没有结皮分布.研究表明地衣-藻结皮是生物土壤结皮的先驱类型,其先于苔藓结皮发育,苔藓结皮是生物土壤结皮发育的成熟阶段[27 ~29 ] .地衣喜少干扰、相对稳定的土壤,其发育还受景观稳定性制约,可作为生态环境变化的指示[6 ] .苔藓结皮仅分布在防护体系防护距离较大,植被带建立较早的区域,表明该区域地表稳定性很好,非常利于结皮发育成熟.地衣- 藻结皮在防护体系内广泛分布,表明这些区域地表处于相对稳定状态,为结皮发育提供了最基本的地表稳定条件.苔藓结皮发育区域,苔藓结皮和地衣- 藻结皮分布在固定沙丘的不同地貌部位,这是生物对环境条件适应的一种生态表现[30 ] .研究证实生物土壤结皮可以显著提高地表的抗风蚀能力,苔藓结皮的抗风蚀能力明显高于地衣- 藻结皮[31 ] ,综上所述,两种生物土壤结皮在防护体系内的差异性分布特征指示了防护体系内地表抗风蚀能力和风沙环境稳定性的空间差异,即沿断面地表稳定性逐渐增强,地势较低和平缓的沙丘地貌部位地表相对更为稳定. ...
... 防护体系内表层沉积物的粒度变化和生物土壤结皮的空间差异表明,沙区风沙环境变化主导着土壤形成发育的方向和速度.防护体系(特别是其中的人工植被和草方格)营造沉积主导的风沙环境,为远距离输送的悬移细颗粒的沉降和富集提供了有利条件,后者则为流沙表面生物土壤结皮的形成发育提供了极其重要的物质基础.粘粒含量是影响土壤肥力的主要因素之一[32 ] ,结皮层中粘粒含量明显提高,既证实沙尘沉降与富集主导的风沙环境的形成,也预示土壤肥力的逐步提高和土壤发育环境的改善.本文结果显示,苔藓结皮和地衣- 藻结皮在沙丘不同地貌部位的分布格局,以及结皮层粘粒含量的变化特征,都与王雪芹等[30 ] 在古尔班通古特沙漠和段争虎等[33 ] 在腾格里沙漠的相关研究结果类似,可见具有普遍意义的是,流沙地区风沙环境性质的改变及其程度和沙丘地形的起伏,是生物土壤结皮形成发育和空间分异的根本原因,而结皮沉积物粒度特征的水平差异性和剖面变化则忠实记录和指示风沙环境的时空变化. ...
古尔班通古特沙漠生物结皮对地表风蚀作用影响的风洞实验
1
2004
... 防护体系前段是风沙活动由强烈风蚀和堆积向沉积为主转变的过渡区域,地表的稳定性不能满足结皮发育,因而没有结皮分布.研究表明地衣-藻结皮是生物土壤结皮的先驱类型,其先于苔藓结皮发育,苔藓结皮是生物土壤结皮发育的成熟阶段[27 ~29 ] .地衣喜少干扰、相对稳定的土壤,其发育还受景观稳定性制约,可作为生态环境变化的指示[6 ] .苔藓结皮仅分布在防护体系防护距离较大,植被带建立较早的区域,表明该区域地表稳定性很好,非常利于结皮发育成熟.地衣- 藻结皮在防护体系内广泛分布,表明这些区域地表处于相对稳定状态,为结皮发育提供了最基本的地表稳定条件.苔藓结皮发育区域,苔藓结皮和地衣- 藻结皮分布在固定沙丘的不同地貌部位,这是生物对环境条件适应的一种生态表现[30 ] .研究证实生物土壤结皮可以显著提高地表的抗风蚀能力,苔藓结皮的抗风蚀能力明显高于地衣- 藻结皮[31 ] ,综上所述,两种生物土壤结皮在防护体系内的差异性分布特征指示了防护体系内地表抗风蚀能力和风沙环境稳定性的空间差异,即沿断面地表稳定性逐渐增强,地势较低和平缓的沙丘地貌部位地表相对更为稳定. ...
沙盖黄土丘陵坡地土壤理化特性随地形变化规律研究
1
2011
... 防护体系内表层沉积物的粒度变化和生物土壤结皮的空间差异表明,沙区风沙环境变化主导着土壤形成发育的方向和速度.防护体系(特别是其中的人工植被和草方格)营造沉积主导的风沙环境,为远距离输送的悬移细颗粒的沉降和富集提供了有利条件,后者则为流沙表面生物土壤结皮的形成发育提供了极其重要的物质基础.粘粒含量是影响土壤肥力的主要因素之一[32 ] ,结皮层中粘粒含量明显提高,既证实沙尘沉降与富集主导的风沙环境的形成,也预示土壤肥力的逐步提高和土壤发育环境的改善.本文结果显示,苔藓结皮和地衣- 藻结皮在沙丘不同地貌部位的分布格局,以及结皮层粘粒含量的变化特征,都与王雪芹等[30 ] 在古尔班通古特沙漠和段争虎等[33 ] 在腾格里沙漠的相关研究结果类似,可见具有普遍意义的是,流沙地区风沙环境性质的改变及其程度和沙丘地形的起伏,是生物土壤结皮形成发育和空间分异的根本原因,而结皮沉积物粒度特征的水平差异性和剖面变化则忠实记录和指示风沙环境的时空变化. ...
Evolution of soil properties on stabilized sands in the Tengger Desert, China
1
2004
... 防护体系内表层沉积物的粒度变化和生物土壤结皮的空间差异表明,沙区风沙环境变化主导着土壤形成发育的方向和速度.防护体系(特别是其中的人工植被和草方格)营造沉积主导的风沙环境,为远距离输送的悬移细颗粒的沉降和富集提供了有利条件,后者则为流沙表面生物土壤结皮的形成发育提供了极其重要的物质基础.粘粒含量是影响土壤肥力的主要因素之一[32 ] ,结皮层中粘粒含量明显提高,既证实沙尘沉降与富集主导的风沙环境的形成,也预示土壤肥力的逐步提高和土壤发育环境的改善.本文结果显示,苔藓结皮和地衣- 藻结皮在沙丘不同地貌部位的分布格局,以及结皮层粘粒含量的变化特征,都与王雪芹等[30 ] 在古尔班通古特沙漠和段争虎等[33 ] 在腾格里沙漠的相关研究结果类似,可见具有普遍意义的是,流沙地区风沙环境性质的改变及其程度和沙丘地形的起伏,是生物土壤结皮形成发育和空间分异的根本原因,而结皮沉积物粒度特征的水平差异性和剖面变化则忠实记录和指示风沙环境的时空变化. ...
 , 张春来
, 张春来

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}