薛红喜 , 李琪, 黄瑜
, 李琪, 黄瑜
XUE Hong-xi, LI Qi, HUANG Yu
中图分类号: S812
文献标识码: A
文章编号: 1000-0690(2014)11-1385-06
通讯作者:
收稿日期: 2013-08-12
修回日期: 2013-11-10
网络出版日期: 2014-11-10
版权声明: 2014 《地理科学》编辑部 本文是开放获取期刊文献,在以下情况下可以自由使用:学术研究、学术交流、科研教学等,但不允许用于商业目的.
基金资助:
作者简介:
作者简介:薛红喜(1978-),男,山西河津人,博士,高级工程师,主要研究方向为农业气象、生态气象。E-mail:xue-hongxi@163.com
展开
摘要
利用涡度相关法对克氏针茅草地生态系统的生态系统初级生产力(GEP)、生态系统呼吸(RECO)和净生态系统碳交换(NEE)进行观测,探讨土壤环境因子对GEP、RECO、NEE的影响。结果表明:2009~2010年,克氏针茅草地生态系统GEP、RECO、NEE都呈现出了明显的峰值变化规律,2010年表现出了更强的固碳能力。土壤环境因子与克氏针茅草地生态系统GEP、RECO、NEE都有极显著的相关性,土壤温度对碳通量组分的影响程度要高于土壤水分;土壤环境因子对GEP、RECO的影响程度要高于对NEE的影响。
关键词:
Abstract
Based on the eddy covariance technique, the variations of the gross ecosystem productivity (GEP), the ecosystem respiration (RECO) and the net ecosystem carbon exchange (NEE) over Stipa krylovii ecosystem were measured in 2009-2010, and the impaction of the soil environmental factors on the GEP, RECO and NEE. The results showed that the GEP, RECO and NEE over the Stipa krylovii ecosystem showed obvious peak change rule in 2009-2010, and the Stipa krylovii ecosystem had the stronger ability of carbon fixation in 2010. There had very significant relationship between the soil environmental factors and the GEP, RECO and NEE over the Stipa krylovii ecosystem. The soil temperature had more profound influence on the carbon flux components of the Stipa krylovii ecosystem than the soil moisture did, and the soil environmental factors had more profound influence on the GEP and RECO than on the NEE.
Keywords:
前 言
碳排放问题是气候变化领域内的研究热点之一[1],草地生态系统几乎覆盖了陆地面积的20%,其净初级生产力约占全球陆地生物区净初级生产力的1/3,在地球碳循环过程中有非常重要的作用[2,3]。与其它陆地生态系统相比,草地生态系统生产力的年际变幅更大,难以确定其碳源汇功能。因此,研究草地生态系统碳循环过程和机理对研究陆地碳循环至关重要[4]。土壤是影响生态系统碳通量变化最直接的因素[5~7],研究土壤温度和水分对草地生态系统碳通量变化的影响,找出环境因子与碳通量之间的关系,对研究草地生态系统碳通量的特征及其对气候变化的影响和响应有重要的意义。
克氏针茅草原是亚洲中部典型的草原代表区系,中心分布区位于蒙古高原,向西与荒漠草原亚带相连,反映出更加旱生的特点。克氏针茅草原所处区域具有干旱与炎热同步的水热变化规律,而土壤环境因子可以反映出这种特点。学者对克氏针茅草地生态系统碳通量变化的驱动机制已有研究[8~10]。但由于观测时间短,数据积累不够,因此其碳通量变化的驱动机制还存在较大的不确定性,还需通过长期观测,为正确认识克氏针茅草地生态系统的碳源汇功能提供基础资料。本研究通过整理分析2009~2010年克氏针茅草地生态系统日尺度上的碳通量变化特征和土壤环境因子的特征,探讨土壤环境因子和克氏针茅草地生态系统碳通量之间的关系,为深入认识克氏针茅草地生态系统碳通量的环境控制机制提供科学依据。
研究区位于内蒙古自治区锡林浩特市东北24 km处(44°08′31″N,116°18′45″E),海拔高度大概1 000 m,下垫面开阔平坦。研究区属温带半干旱草地气候,年平均气温为2.6℃,1月均温-22.0℃,7月均温18.3℃;日照充足,年均日照总时数约为2 600 h;春、秋、冬3季干燥少雨,降水主要集中在6~9月,年降水量约为290 mm。土壤为典型的栗钙土。研究区处于内蒙古典型的克氏针茅草地区,群落以克氏针茅和羊草为优势草种,伴生有糙隐子草、冰草等禾本科草类以及菊科的冷蒿等。
本研究使用涡度相关系统观测克氏针茅草地生态系统碳通量。开路式涡度相关系统由三维超声风速仪和开路式CO2/H2O分析仪构成,高度为4 m。使用铂电阻地温仪测定10 cm土壤处的土壤温度;使用土壤水分传感器测定10 cm深处的土壤水分含量。通量采集频率为10 Hz,土壤环境要素采样频率为0.5 Hz,所有的数据由特定的数据采集器按30 min步长计算并存储平均值。按照微气象学符号协定,碳通量的负值表示碳吸收,正值表示碳排放。
选取2009~2010年两年的碳通量观测数据,进行坐标旋转、WPL校正等处理,并对由系统故障、供电、下雨或检修等因素造成的丢失数据进行了剔除。采用查表法和日平均变化法(对无法用查表法以及非线性回归法进行插补的缺失数据,采用此方法)对通量数据进行插补,并按30 min步长计算平均值,再分别计算日统计数据[11]。
利用Excel和SPSS等软件研究土壤环境因子和克氏针茅草地生态系统碳通量变化的相关关系,从而深入理解克氏针茅草地生态系统碳通量变化的驱动机制。
生态系统净碳通量(NEE,值表示为NNEE)的定义式为[12~14]:
式中,上划线表示某时间段内的平均值,撇号表示脉动,即瞬时值与平均值的偏差。等号右边第1项为CO2湍流通量,即单位时间垂直方向上通过单位截面积的CO2的量,
呼吸作用是夜间生态系统植被与大气间的NEE通量的唯一来源,生态系统夜间的RECO(值表示为Reco)可用Lloyd-Taylor方程进行插补:
式中,Reco,ref为参考温度下的生态系统夜间呼吸;Tref为参考温度,取值为283.15 K;Tk为夜间绝对温度,单位为K;T0为温度试验常数,取值为227.13 K,E0为活化能,取值为308.56 K。生态系统白天的呼吸是根据夜间生态系统呼吸数据建立的函数关系外延到白天后获得的。
由于白天植被与大气之间的NEE是生态系统初级生产力(GEP,值表示为GGEP)和生态系统呼吸(RECO)的总和,因此GGEP就可定义为:
GGEP=NNEE- Reco (3)
图1显示了2009和2010年克氏针茅草地生态系统GEP日总值的变化情况。图1看出,2009、2010年GEP日总值变化都呈现出了不明显的“W”型波峰,其中第一个波峰都比较明显,第二个波峰不明显,且明显小于第一个波峰,但第二个波峰的持续时间要更长一些。2009年的GEP最小值[-2.88 g/(m2·d)]明显要大于2010年的[-4.47 g/(m2·d)],说明2010年克氏针茅草地生态系统的生长更加旺盛。
图2显示,2009和2010年Reco日总值变化均呈明显的倒“U”型。Reco在两年中的变化规律基本一致,即Reco先随时间变化增大,大概在7月下旬出现最大值,然后随着时间推移而逐渐减小。2009年Reco最大值为1.34 g/(m2·d),最小值为0.10 g/(m2·d);2010年Reco最大值为1.52 g/(m2·d),最小值为0.21 g/(m2·d),说明2010年克氏针茅草地生态系统的呼吸作用更旺盛。
从图3可以看出2009和2010年NEE日总值也呈不明显的“W”型波峰,与GEP的变化规律基本相同。波峰出现的时间与GEP的峰值出现时间也基本一致。从NEE的波动范围来看,2009年的波动幅度较小[最大最小值分别为0.81和-1.84 g/(m2·d)],2010年NEE值的波动幅度[最大最小值分别为1.2和-3.64 g/(m2·d)]明显大于2009年,说明2010年克氏针茅草地生态系统的碳交换活动更加活跃。
从图4可知,2009~2010年克氏针茅草地生态系统的土壤温度日变化趋势基本相同,都呈明显的倒“U”型,且增减过程都较平缓。2010年的土壤平均温度(4.9℃)低于2009年(5.1℃),但在克氏针茅的主要生长季(5~10月),则是2010年的土壤平均温度(16.9℃)高于2009年(16.1℃)。2009~2010年克氏针茅草地生态系统的土壤水分含量在正常范围内变化,2009年的土壤水分含量只出现了一个峰值,而2010年的土壤水分含量出现多个峰值,且2010年土壤水分含量的平均值(0.12 m3/m3)高于2009年(0.10 m3/m3),因此2010年的土壤水分条件要好于2009年。从整体来看,2010年克氏针茅草地生态系统的土壤温、水环境要好于2009年,而对应的2010年的克氏针茅草地生态系统的GGEP、Reco、NNEE的变幅都要大于2009年,2010年克氏针茅草地生态系统显示出了更旺盛的生长特征。
图5显示了2009~2010年土壤温度和水分对克氏针茅草地生态系统GEP的影响。从图中可以看出,GEP与土壤温度和水分都呈二次方关系,显著性检验结果表明,GEP与土壤温度和水分都呈极显著(p<0.01,下同)的相关关系(相关系数分别为0.72和0.66)。从拟合的趋势线可以看出,克氏针茅草地生态系统GEP绝对值随土壤温度的升高以及土壤水分含量的增加而增大,说明土壤温度和水分提高可以促进克氏针茅草地生态系统的初级生产力活动。另外从两个方程的决定系数来看,土壤温度和土壤水分分别可以解释克氏针茅草地生态系统GEP变化的52%和44%,土壤温度对GEP的影响略大一些。
图6显示了2009~2010年土壤温度和水分对克氏针茅草地生态系统Reco的影响。从图中可以看出,克氏针茅草地生态系统呼吸作用与土壤温度和土壤水分都呈极显著的相关关系(相关系数分别为0.91和0.70),且Reco随着土壤温度和水分的升高都以指数方式增长。从方程的决定系数来看,土壤温度可以解释克氏针茅草地生态系统Reco83%的变化量,远高于土壤水分对Reco的影响,说明土壤温度是影响克氏针茅草地生态系统呼吸作用的关键因子。
图4 2009~2010年土壤温度和水分的日变化
Fig.4 Changes of soil temperature and moisture in 2009-2010
图5 2009~2010年土壤温度和水分对GGEP的影响
Fig.5 The effect of soil temperature and moisture on GGEP in 2009-2010
图6 2009~2010年土壤温度和水分对Reco的影响
Fig.6 The effect of soil temperature and moisture on Reco in 2009-2010
图7显示2009~2010年土壤温度和水分对克氏针茅草地生态系统NEE的影响。从图中可以看出,NEE与土壤温度和水分都呈极显著的相关关系(相关系数都为0.43),随着土壤温度和水分的升高,NEE的绝对值有增大的趋势。但从拟合方程的决定系数来看,土壤温度和水分对克氏针茅草地生态系统NEE的影响较小,只能解释NEE18%左右的变化量。
已有的研究结果表明,土壤环境条件是影响草地生态系统碳通量的重要因素,其中土壤温度和水分是草地生态系统碳通量不同组分时间变化的重要驱动因子[15~20]。本文的研究结果也表明,10 cm处的土壤温度和水分对克氏针茅草地生态系统GEP、RECO和NEE都有一定的影响,且都呈现出了极显著的相关关系,土壤环境因子是克氏针茅草地生态系统碳通量的重要驱动因子,这与其他研究结果是相似的。
本文的结果同时表明,土壤温度和水分对克氏针茅草地生态系统碳通量的3个组分的影响程度是不一样的。从拟合方程的决定系数来看,土壤温度和水分对GEP和RECO的影响远大于对NEE的影响。从计算公式上可以看出,NEE是GEP和RECO的和,因此GEP和RECO可以看作是碳通量的直接组分,而NEE是碳通量的间接组分。从这个角度来看,土壤温度和水分对克氏针茅草地生态系统的初级生产力和生态系统呼吸这两个直接碳通量组分的影响程度更高,而对NEE这个间接的碳通量组分的影响程度较低。
研究表明,土壤水分会影响到土壤温度对草地生态系统碳通量的作用[20]。本文的研究结果也表明(表1),在土壤水分条件较好的2010年,土壤
图7 2009~2010年土壤温度和水分对NNEE的影响
Fig.7 The effect of soil temperature and moisture on NNEE in 2009-2010
温度与GEP、RECO和NEE拟合方程的决定系数(分别为0.57、0.85和0.27)都要高于2009年(分别为0.48、0.80和0.10)。王云龙[7]的研究结果也表明,水分条件较好的2004年,克氏针茅草地生态系统RECO与土壤温度之间的相关系数要高于水分条件较差的2005和2006年。这一结果说明在土壤水分充足时,土壤温度能够更好地解释克氏针茅草地生态系统碳通量的变化。克氏针茅草地生态系统为半干旱生态系统,其所处的区域降雨较少,土壤水分状况会限制土壤温度对克氏针茅草地生态系统固碳能力的影响,因此土壤水分是克氏针茅草地生态系统碳源汇转换的关键因子。
flux under different soil moisture conditions
| 2009年 | 2010年 | |
|---|---|---|
| GGEP | y=-0.0003x2-0.0269x-0.2415 R2=0.4795 | y=-0.001x2-0.0302x-0.1981 R2=0.5725 |
| Reco | y=0.448e0.0342x R2=0.8041 | y=0.3997e0.0365x R2=0.851 |
| NNEE | y=9E-05x2-0.0111x+0.1963 R2=0.1025 | y=-0.0007x2-0.0139x+0.2139 R2=0.2705 |
碳通量及其驱动机制是全球碳循环研究的焦点[13,21],本文利用两年的观测数据对克氏针茅草地生态系统进行了研究,其结论还存在一定的不确定性,还需要多年观测数据的积累才能对碳通量的驱动机制有清晰认识;另外不同的生态系统类型对环境要素的响应机制是不同的,还需要建立更多的观测站点并进行长期研究,才能对全球碳循环机制有更全面的了解。
1) 2009~2010年,克氏针茅草地生态系统GEP、RECO、NEE都呈现出了明显的峰值变化规律,2010年表现出了更加旺盛的初级生产力以及系统呼吸能力,其固碳能力也更强。2010年的土壤温度和水分条件也要好于2009年。
2) 2009~2010年,土壤环境因子与克氏针茅草地生态系统GEP、RECO、NEE都有极显著的相关性,土壤温度对克氏针茅草地生态系统碳通量组分的影响程度要高于土壤水分;土壤环境因子对克氏针茅草地生态系统GEP、RECO的影响程度要高于对NEE的影响。
3) 土壤温度和土壤水分都是克氏针茅草地生态系统碳通量变化的重要驱动因子,土壤水分条件会限制土壤温度对碳通量的影响程度,因此土壤水分是克氏针茅草地生态系统碳源汇转换的关键因子。
The authors have declared that no competing interests exist.
| [1] |
干旱区不同土地利用方式下土壤呼吸日变化差异及影响因素 [J]. |
| [2] |
草地生态系碳循环研究评述 [J]. |
| [3] |
青海省草地资源净初级生产力遥感监测 [J]. |
| [4] |
McInnes K J,et al.Carbon dioxide exchange in a subtropical, mixed C3/C4 grassland on the Edwards Plateau,Texas [J]. |
| [5] |
Carbon dioxide exchange between the atmosphere and an alpine meadow ecosystem on the Qinghai-Tibetan Plateau,China [J]. |
| [6] |
Depression of net ecosystem CO2 exchange in semi-arid Leymus chinensis steppe and alpine shrub [J].
|
| [7] |
Seasonal variation in carbon dioxide exchange over a Mediterranean annual grassland in California [J]. |
| [8] |
土壤温度和水分对克氏针茅草原生态系统碳通量的影响初探 [J]. |
| [9] |
克氏针茅草地的碳通量与碳收支[D] .
|
| [10] |
温度对克氏针茅草原生态系统生长季碳通量的影响 [J]. |
| [11] |
“Turning point air temperature” for alpine meadow ecosystem CO2 exchange on the Qinghai-Tibetan Plateau [J]. |
| [12] |
Recent progress and future directions of ChinaFLUX [J]. |
| [13] |
|
| [14] |
中国北方针叶林生长季碳交换及其调控机制 [J]. |
| [15] |
草地生态系统碳蓄积的研究进展 [J]. |
| [16] |
青藏高原高寒草甸生态系统净二氧化碳交换量特征 [J].
|
| [17] |
青藏高原矮嵩草草甸和金露梅灌丛草甸CO2通量变化与环境因子的关系 [J].
|
| [18] |
极端干旱条件下锡林河流域羊草草地净生态系统碳交换特征 [J].
|
| [19] |
黄土丘陵区天然草地CO2通量研究[D] .
|
| [20] |
青藏高原高寒灌丛生长季和非生长季CO2通量分析 [J]. |
| [21] |
黑河中游不同土地利用类型下土壤碳储量及其空间变化 [J]. |

/
| 〈 |
|
〉 |



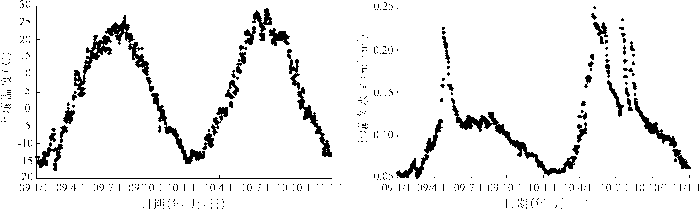



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}