许振文 , 王晓东
, 王晓东
XU Zhen-wen, WANG Xiao-dong
中图分类号: K903
文献标识码: A
文章编号: 1000-0690(2014)06-0748-09
通讯作者:
收稿日期: 2013-12-3
修回日期: 2014-02-19
网络出版日期: 2014-06-10
版权声明: 2014 《地理科学》编辑部 本文是开放获取期刊文献,在以下情况下可以自由使用:学术研究、学术交流、科研教学等,但不允许用于商业目的.
基金资助:
作者简介:
作者简介:许振文(1969-),男,吉林长春人,副教授。主要从事自然地理研究。E-mail: wangxd219@nenu.edu.cn
展开
摘要
以长白山北坡林线长白落叶松(Larix olgensis)和岳桦(Betula ermanii)为研究对象,通过野外样方调查、树轮技术分析,采用趋势线模型拟和、单因素方差等统计方法从各种参数分析长白落叶松和岳桦径向树高生长的对比关系并揭示2个种群动态对环境适应性的对比差异。结果显示:垫状生长形的长白落叶松海拔最高(均值:2 017 m),其次为旗形树(2 005 m),正常形态的海拔最低(1 992 m),所在生境差异显著(p < 0.05)。正常形态的长白落叶松生长速度大于岳桦(树高方程斜率:长白落叶松0.750 9>岳桦0.253 6),随着旗形树的出现,速度相似(斜率:长白落叶松0.375 4>岳桦0.253 6),变成垫状后,岳桦占优。长白落叶松受到压制种群扩张速度不如岳桦,长白山林线上侵表现为以岳桦种群为主力,长白落叶松仍是岳桦主要的伴生树种。
关键词:
Abstract
The aim of this article is to understand the different responses between Laris olgensis and Betula ermanii to environment adaptability on north slopes treeline of the Changbai Mountains. The alpine treeline is regarded as highly sensitive and a potential indicator of environmental change and it has thus become one of the hotspots of environmental change research. Most researches on treeline dynamics have been conducted at a population scale, and there have been comparatively few studies of treeline dynamics at multi-population. In this study, treeline dynamics were analyzed at two populations in the Changbai Mountains by examining the change in relationship between two population’s dynamics and the environment. The treeline of the northern slope of the Changbai Mountains was selected as the study area. Data source was obtained by field investigation and tree-ring analysis. The altitude differences of three growth forms (general tree, flag tree and cushion) of Laris olgensis were revealed by half-peak width, mean ± standard deviation, and One-way ANOVA. DBH (diameter at breast height) of each tree (Laris olgensis and Betula ermanii) was measured by measuring tape (precision: 0.1 cm) and 142 cores were taken from the living trees in plots. The ages of the sampling trees were known using tree-ring and the age-class (DBH) regressions were created to estimate ages of all trees in the plots. The annual stem recruitments of Laris olgensis and Betula ermanii populations were calculated based on field investigations. The difference of growth rates between Laris olgensis and Betula ermanii was understood by assessing differences in the slopes of trend lines and paired-samples T test. The different responses to environmental change between Laris olgensis and Betula ermanii were evaluated using mathematical models. The results showed that cushion growth form of Laris olgensis situated in the highest habitat, followed by flag tree and general tree in the lowest habitat. There were significant differences among three habitats of growth forms of Laris olgensis (p<0.05). The growth velocity of diameter and height growth of Laris olgensis with general growth form was higher than that of Betula ermanii. The difference was not significant between Laris olgensis and Betula ermanii when Laris olgensis changed flag tree. However, Laris olgensis was lower than Betula ermanii when it changed cushion growth form. Population expansions of Laris olgensis and Betula ermanii were very obvious as temperature increase. Expension velocity of Laris olgensis was lower than that of Betula ermanii because it was pressed by Betula ermanii. The growth velocity of all growth forms of Laris olgensis was lower than that of Betula ermanii when tree ages were less than 13 years. Treeline shift was caused mainly by population expansion of Betula ermanii. Laris olgensis was still main accompanying tree specie of Betula ermanii. However, Laris olgensis may become pure forest when Betula ermanii was cut down in quantity.
Keywords:
长白山北坡岳桦(Betula ermanii)林是以单一乔木树种为主的林线植被[1],是东亚地区保存最为完整的岳桦林带[2]。除有水土保持、涵养水源等生态功能外[3],也是长白山旅游区的重要景点[4],具有独特的旅游价值[5]。但是在景区开发与经济建设中岳桦林受到一定的人为破坏,同时,岳桦林作为先锋物种有被针叶林替代的趋势[6]。另外野外观测时发现在岳桦林中混有长白落叶松[7],即使对森林生长受限的林线也有这种树的分布[8],据此现象推测长白落叶松可能拥有替代岳桦的机会[9]。
在一些探讨岳桦动态的研究中[10,11]涉及到长白落叶松(Larix olgensis)的研究,如应用生态重要值讨论长白落叶松的地位[12],论述气候变化与长白落叶松生长的关系[13],对立地条件,林分密度,立木更新等生长指数进行讨论[14]。在应用LINKAGES模型分析主要乔木树种对气候变化的响应中发现长白落叶松呈扩张态势[15]。在各地林线研究时发现针叶树出现生长形态变化的现象[16~ 18]。在长白山岳桦林线,长白落叶松存在与正常生长形态不同的旗形与垫状树。
虽然许多研究都涉及到岳桦林内的长白落叶松,但是对长白落叶松与岳桦的相互关系分析不足,尤其是其生长形态演变的分布规律以及这种演变特征与环境之间的关系更是缺乏研究。所以本文利用野外样方调查与树轮技术相结合,应用方差分析、数学模型等常规统计方法对正常与特殊生长形态的长白落叶松和岳桦径向树高生长速度进行对比,揭示2种乔木种群的发展态势和对环境(以气候为代表)适应的方式,使生态系统对全球变化响应的研究从单一种群向多个种群乃至生态系统的变动机理分析提供一种新的思考方向。
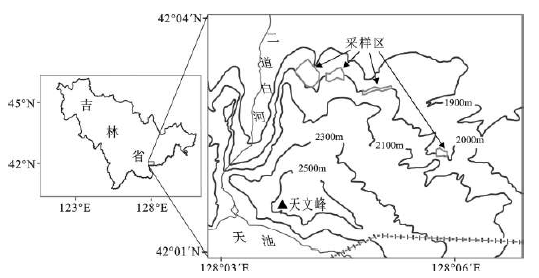
长白山北坡林线海拔为1 900~2 000 m左右[19],林线生境风大寒冷[20],岳桦由于其较强的抗寒、抗风而成为长白山森林分布的上限树种[21],同时与长白落叶松、东北赤扬(Alnusmandshurica)等少量乔木混生[22]。树线乔木层植被为岳桦纯林和岳桦-长白落叶松混交林两种类型[23],林线处岳桦以斑块状镶嵌分布于高山苔原带,并逐渐向上扩展[2,24]。研究区内林线型为岳桦-长白落叶松混交林,长白落叶松除正常生长形态外,一些树木的生长形态出现变化,如旗形树,即树木的一侧枝叶完全消失,只在另外对应一侧生长,使树木的外形呈现旗形。另外一种变形是主干没有长成,而是一些支干在地面呈现匍匐生长状态,在外貌显示垫状形态。取样区地理位置为42°2′27″N~ 42°3′14″ N,128°4′35″E~ 128°5′53″ E,是长白山森林-苔原过渡带典型地段(图 1)。
图 1 长白山北坡林线采样区
Fig. 1 Sampling plots within the treeline on the north-facing slope of the Changbai Mountains
2009年9月采用样方调查法在研究区内共取4个样地,即考虑地形分异对气候等环境因子的影响,只选择在局部地形区中,坡向为正北的地形区为样地,每个样地水平宽度(与等高线垂直)大约120~ 250 m,长度(沿等高线方向)大约40~ 210 m(表 1)(图 1)。在每个样地内水平方向上大约每间隔40 m左右设置一样带,即从最前缘(海拔最高处)(树岛)沿等高线向下(自高而低)一直到海拔较低的郁闭林(郁闭度>0.2),这样形成宽为10 m与等高线垂直的长条型样带。4个样地一共得到13条样带,样带的长度不同,分别从40 m到210 m(表 1)。在每一个样带内设置样方,即从样带最前缘(树岛)连续取样,一直到样带最下方(郁闭林),这些每一样带内拥有多个紧密相连的样方。样方大小为10 m×10 m,共得到153个样方(图 1)。在得到的样方内选择既有长白落叶松又有岳桦的57个样方进行立木调查,测量内容有样方的海拔高度、地理位置、长白落叶松和岳桦的个体数量、胸径(距地面1.2 m)、树高大小,共观测到1 898棵岳桦和334棵长白落叶松,在这些落叶松中有20棵垫状树;48棵旗形树;266棵正常形态的树。用GPS测定样地的经纬度和海拔,胸径用卷尺测量(精确到0.1 cm),树高不超过2 m用直接测量法(带刻度的标杆),对超过2 m的树高用测绳测量,对于个别高大树木用全站仪测量,精度为0.1 m (表 1)。
表 1 样地的基本特征
Table 1 Plots characters of the alpine treeline on the northern slope of the Changbai Mountains
| 样地 | 海拔(m) | 样带条数 | 样带长度(m) | 样方个数(个) | 样方总数(个) | 取样方总数(个) | 岳桦棵数(棵) | 落叶松棵数(棵) |
|---|---|---|---|---|---|---|---|---|
| 1 | 1968~2032 | 5 | 110~210 | 11~21 | 75 | 22 | 1057 | 74 |
| 2 | 1994~2024 | 3 | 90~100 | 9~10 | 28 | 8 | 251 | 32 |
| 3 | 1970~1989 | 2 | 40~50 | 4~5 | 9 | 4 | 170 | 27 |
| 4 | 1979~2024 | 3 | 90~180 | 9~18 | 41 | 23 | 420 | 201 |
由于岳桦和长白落叶松都适合做树木年轮分析[25],在长白山也有以岳桦为树种进行轮宽研究的先例[26],所以在测量的树木中按照径级分布选择一些长白落叶松和岳桦个体做树轮分析,树芯取样的方法是在胸径(距地面1.2 m)处用生长锥取2个树芯,小径级在距地表0.3 m的树干处取样,共得到142个样品。经干燥打磨后在仪器(LA-S型植物年轮分析系统:2亿像素)上交叉定年[27],排除低质量的38个后获取104样品年轮数据。按照径级(胸径)、树高与对应的年轮数据(树龄)的关系,分别建立岳桦和长白落叶松的径级(胸径)-年龄(树龄)、径级(胸径)-树高生长的数学模型。同样利用得到的年轮数据分别建立岳桦和长白落叶松的年龄(树龄)-径级(胸径)、年龄(树龄)-树高的数学模型,在众多数学模型中取拟和度最好的指数模型(公式1、2、3、4)。依据长白落叶松大约百年达到过熟林的情况[28],即超过百年时树高和径级可能停止生长,所以只将方程的年龄推到100 a。
长白落叶松:
Y = 0.4442 X1.7538 (R2 = 0.966, P<0.001) (1)
岳桦:
Y = 0.0425 X 1.3455 (R2 = 0.949, P<0.001) (2)
Y 是径级(cm),X 是年龄(a)。
长白落叶松:
H = 0.0983 X1.2126 (R2 = 0.876, P<0.001) (3)
岳桦:
H = 0.4165 X0.5852 (R2 = 0.937, P<0.001) (4)
H 是树高(m),X 是年龄(a)。
利用徐文铎[29]、吴正方[30]采用的半峰宽(即通过峰高的中点作平行于峰底的直线,此直线与峰两侧相交2点之间的距离)和均值
依据径级结构反映年龄结构[31]的原理,分析长白落叶松和岳桦种群径级结构揭示种群动态及其对比关系。具体分析是先按照长白落叶松生长形态的差异将长白落叶松和岳桦种群动态的对比分3组进行,即把所有生长形态为垫状的长白落叶松样方内的长白落叶松和对应样方内的岳桦分别按照径级大小进行统计,旗形和正常形态的长白落叶松也按照这样方法进行分组统计,最后得到3组长白落叶松和岳桦种群径级分布的对比数据。
径级结构分布的统计是将样方调查得到的树木棵数按照径级(胸径)大小(四舍五入取整数)进行统计,根据树木生长规律把径级从大向小排布,计算每一径级上的树木棵数。然后依据年龄(树龄)-径级(胸径)方程递推长白山林线气温波动过程中每一高温期前后径级结构的数量分布,再将径级分布规律做直线趋势拟和,依据直线拟和方程斜率值的大小判断种群动态及其2个种群发展态势的对比。
3.1.1 不同生长形生境海拔的分化
野外样方调查显示3种生长形在海拔分布上有一定差异,从全范围来看,呈现小范围的混合交错。两种适宜范围的分析方法结果都显示:垫状生长形海拔最高(均值
表 2 不同生长形态的长白落叶松生存环境的海拔和适宜范围
Table 2 Attitude and optimum spatial scopes of three growth forms of Larix olgensis
| 生长 状态 | 分布范围 (m) | 适宜范围(m)(均值 | 适宜范围(m)(半峰宽) |
|---|---|---|---|
| 垫状 | 2000~2032 | 2017 | 2006~ 2028 |
| 旗形 | 1986~ 2024 | 2005 | 1990~ 2021 |
| 正常形 | 1974~ 2019 | 1992 | 1977~ 2006 |
3.1.2 生境海拔的差异程度
长白落叶松所在样方海拔的单因素方差分析结果显示:在空间分布上3种形态存在显著差异(p<0.05),正常生长形态与其它两种形态之间差异极显著(p<0.01)(表 3),显示不同生长形所在空间范围是不同的,由于海拔越高,气温越低,生存环境越恶劣,而三者所在的海拔范围有着明显的差异,体现海拔差异所产生的气候梯度差是长白落叶松形态变化的主要因素。长白落叶松正是通过调整自己的生长方式来实现对不同环境的适应,从而达到扩展生存范围。但是这种变化带来的一个结果是竞争力相应下降,而这种下降又在与林线的建群种-岳桦生境重合的情况下变化的,两种树木生态位的竞争态势必然随着海拔增加,长白落叶松生长形态的变化而相应转变。因为树高与径向生长速度是影响竞争力的关键,所以利用生长速度的差异程度判断两种乔木在不同情况下竞争力强弱的对比态势。
表3 不同生长形长白落叶松的海拔差异
Table 3 Altitude differences among different growth forms of Larix olgensis
| 长白落叶松不同生长形态对比 | 高度均值差(m) | 标准差 | p值 | |
|---|---|---|---|---|
| 旗形 | 正常形 | 13.610* | 4.510 | 0.000 |
| 垫状 | 旗形 | 11.800* | 5.440 | 0.030 |
| 正常形 | 25.410* | 4.170 | 0.000 | |
3.2.1 径向生长的对比分析
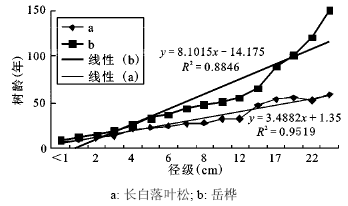
长白落叶松和岳桦的径级(胸径)-年龄生长曲线的趋势线拟和效果较好(R2 > 0.800,p<0.001),可以用于树高生长的分析。从方程参数来看,当长到相同径级时长白落叶松所需时间小于岳桦(斜率:长白落叶松3.488 2<岳桦8.101 5)(图 2),配对样本T检验结果显示当达到相同径级时所用时间有显著差异(p<0.001),说明长白落叶松径向生长速度显著大于岳桦。
图 2 长白落叶松和岳桦的径级-年龄生长曲线
Fig. 2 The growth curve of diameter-year of Larix olgensis and Betula ermanii
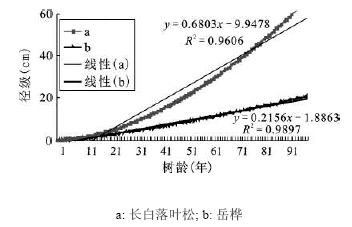
从拟合的树龄-径级(胸径)生长曲线(图 3)可以看出长白落叶松径向生长速度大于岳桦,进一步说明从径向生长速度看长白落叶松竞争力明显大于岳桦。但是从趋势线的变化来看在树龄在13 a左右长白落叶松径向生长速度开始超过岳桦(图 3),表明在幼苗幼树时期岳桦的竞争力大于长白落叶松,即长白落叶松径向生长潜力大于岳桦得渡过幼苗幼树期才能实现。
图 3 长白落叶松和岳桦的树龄-径级曲线
Fig. 3 The growth curve of year-diameter of Larix olgensis and Betula ermanii
3.2.2 树高生长的对比分析
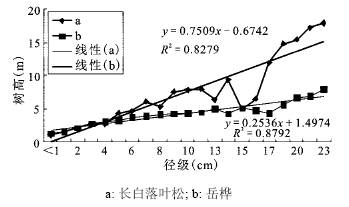
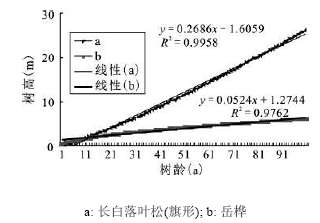
长白落叶松和岳桦径级(胸径)-树高生长曲线的趋势线拟和效果较好(R2 > 0.800,p<0.001),从方程参数来看,在相同径级下长白落叶松树高大于岳桦(斜率:长白落叶松0.750 9>岳桦0.253 6)(图 4)。配对样本T检验结果显示长白落叶松和岳桦在径级相同的情况下树高有显著差异(p < 0.001),说明长白落叶松树高生长速度显著大于岳桦。
图 4 长白落叶松和岳桦径级(胸径)-树高生长曲线
Fig. 4 The growth curve of diameter-height of Larix olgensis and Betula ermanii
在林线附近,树高达到某一高度后,因为本身因素和气候等因子的影响高度增加缓慢[32]。野外观测发现长白山北坡林线上岳桦树高很少超过8 m。为了更好的进行树龄-树高的拟合对比,除以100 a作为一个极限外,再增加岳桦树高8 m作为增长的极限。从拟合的树龄-树高生长曲线看出长白落叶松树高生长速度大于岳桦(图 5),说明从树高生长速度看长白落叶松竞争力明显大于岳桦。但是从趋势线的变化看树龄在13 a左右长白落叶松树高生长速度开始超过岳桦(图 5),同样表明长白落叶松若将树高生长速度超过岳桦的潜力释放必须度过幼苗幼树期。
图 5 长白落叶松和岳桦树龄-树高曲线
Fig. 5 The growth curve of height-year of Larix olgensis (flag tree) and Betula ermanii
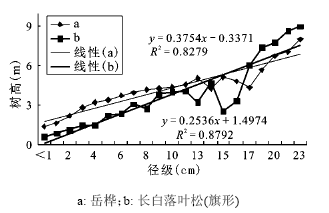
从树高生长速度的对比结果反映岳桦不及长白落叶松,但是野外观测时发现在一定的海拔上,出现旗形树。由于树木一侧没有枝叶,只有利用另外一侧进行光合作用等生理活动,所以推测竞争能力比相对正常生长的树木减少一半。按照这种设想进行比较运算,即将竞争力减半,把旗形树的竞争力(以树高为参照指标)减少一半再进行径级(胸径)-树高(竞争力)的拟和运算。长白落叶松和岳桦生长曲线的趋势线拟和效果较好(R2 > 0.800,p<0.001),从方程参数来看,长白落叶松竞争力接近岳桦(斜率:长白落叶松0.375 4>岳桦0.253 6)(图6),从配对样本T检验结果来看(p = 0.114>0.050),旗形长白落叶松和岳桦的生长过程中竞争力没有显著差异。
图 6 长白落叶松(旗形)和岳桦径级(胸径)-树高生长曲线
Fig. 6 The growth curve of diameter-height of Larix olgensis (flag tree) and Betula ermanii
20世纪50年代以来长白山林线气温在波动上升中出现3次高温期[33],而林线以岳桦为代表的乔木种群对气温上升的响应是种群迅速扩张,个体数量急剧增加[20],长白落叶松也会随着环境趋好种群膨胀,这样对生态位的竞争必然加剧,竞争力的对比在对环境适应性扩张过程中得以显现出来,所以按照种群变动与气温变化的关系将种群径级结构分为3个阶段进行分析,分别判断在每一次高温期前后长白落叶松和岳桦种群径级结构特征并进行对比分析以揭示当环境变化时两者响应方式的差异(表 4)。
表 4 长白落叶松和岳桦种群径级结构趋势线的斜率
Table 4 Trendline slope comparison of diameter-class structure between Larix olgensis and Betula ermanii
| 生长形态 | 阶段 | K | R2 | ||
|---|---|---|---|---|---|
| 长白落叶松 | 岳桦 | 长白落叶松 | 岳桦 | ||
| 垫状 | 第1次高温期前(T1b) | 1.5357 | 0.5091 | ||
| 第1次高温期后(T1a) | 1.6500 | 0.5155 | |||
| 第2次高温期前(T2b) | 1.2485 | 0.4049 | |||
| 第2次高温期后(T2a) | 1.6364 | 0.6291 | |||
| 第3次高温期前(T3b) | 2.1978 | 0.6684 | |||
| 第3次高温期后(T3a) | 5.0980 | 0.5470 | |||
| 正常生长型 | 第1次高温期前(T1b) | 0.2253 | 0.1536 | 0.2809 | 0.3025 |
| 第1次高温期后(T1a) | 0.1395 | 0.2066 | 0.1545 | 0.2901 | |
| 第2次高温期前(T2b) | 0.2617 | 0.2364 | 0.2700 | 0.3474 | |
| 第2次高温期后(T2a) | 0.2286 | 0.4555 | 0.2256 | 0.3695 | |
| 第3次高温期前(T3b) | 0.2150 | 0.6053 | 0.2684 | 0.384 | |
| 第3次高温期后(T3a) | 0.4782 | 3.9300 | 0.3881 | 0.2098 | |
| 旗型 | 第1次高温期前(T1b) | 0.5000 | 0.2632 | ||
| 第1次高温期后(T1a) | 0.8571 | 0.665 | |||
| 第2次高温期前(T2b) | 0.2000 | 0.3571 | 0.1000 | 0.1645 | |
| 第2次高温期后(T2a) | -0.1000 | 0.7000 | 0.0357 | 0.5422 | |
| 第3次高温期前(T3b) | 0.3000 | 0.7000 | 0.2250 | 0.5422 | |
| 第3次高温期后(T3a) | 0.4901 | 4.2110 | 0.4305 | 0.6188 | |
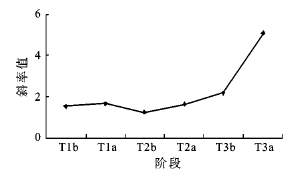
3.3.1 垫状长白落叶松样地内岳桦径级结构的变化
在垫状长白落叶松的样地内,由于垫状落叶松无法确定年龄,只分析岳桦径级结构的变化,不同时期的岳桦径级结构的趋势线斜率呈波状增加,从每一个高温期斜率前后对比来看,高温期后都比高温期前大(表 4,图 7),说明每一个高温期都是种群扩张积极的时期。从几个高温期的斜率增加值看一次比一次大[(0.114 3(KT1a-KT1b)<0.387 9(KT2a-KT2b)<2.900 2(KT3a-KT3b)],与高温期增温幅度逐渐加大的趋势相吻合[33]。不同时期斜率的对比变化进一步证明岳桦种群对气温变化响应明显,气温升高促使岳桦种群不断扩张且与升温幅度密切相连。
图 7 3个高温期前后岳桦径级结构趋势线斜率的变化
Fig. 7 The trendline slope change of diameter-class structure of Betula ermanii on pre- and post-three megathermal periods
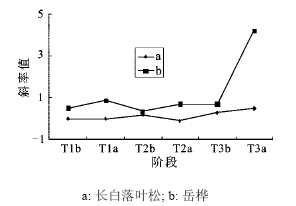
3.3.2 旗形长白落叶松和岳桦径级结构的对比分析
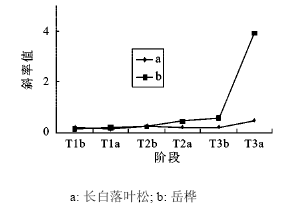
在旗形长白落叶松的样地内,不同时期的岳桦径级结构的趋势线斜率呈波状增加,从每一个高温期斜率前后对比和几个高温期的斜率增加值差异来看,岳桦种群扩张特点与垫状长白落叶松样地内岳桦扩张相似(表 4,图 8)。而旗形长白落叶松则较复杂,在前两个高温期的变化与岳桦变化趋势相反,第三个高温期相似都保持迅速增加态势(表 4,图 8),说明在前2个高温期内由于升温幅度小,在样地内只能共同挤占狭小局部有利的生境,而幼苗阶段岳桦竞争力占优,岳桦种群的膨胀挤压了长白落叶松的扩张,长白落叶松只能在非高温期岳桦种群扩张放缓的情况下加速扩张。由于两者的竞争力相似,这种差异不明显。第三高温期内,由于升温幅度大,两者扩张都向面状地域展开[33],生态位竞争趋缓,两个种群都迅速扩张,但是从两者在这个阶段的斜率大小差异看[(4.211 0(KT3a岳桦)>0.490 1(KT3a长白落叶松)]长白落叶松还有受岳桦挤压的痕迹。
图 8 3个高温期前后旗形长白落叶松和岳桦径级结构趋势线斜率的变化
Fig. 8 The trendline slope change of diameter-class structure of Larix olgensis (flag tree) and Betula ermanii on pre- and post-three megathermal periods
3.3.3 正常形态长白落叶松和岳桦径级结构的对比分析
在正常形态长白落叶松的样地内,不同时期的岳桦径级结构的趋势线斜率呈波状增加,但波动幅度比较小,几乎一直保持增加的态势(表 4,图 9)。长白落叶松也保持增加态势,但是增幅明显小于岳桦,表明受到岳桦种群膨胀的极大挤压。仅在第一个高温期长白落叶松扩张占有优势,在第一个高温期末岳桦扩张的能力就超过长白落叶松,在第二个高温期前虽然长白落叶松扩张能力与岳桦相似,但是在第二高温期后岳桦扩张的能力则明显超过长白落叶松,而且差距越来越大(表 4,图 9)。这个对比差异显示在林线海拔较低处,虽然长白落叶松拥有较强的竞争力,但是由于在幼苗阶段处于弱势,使其在岳桦扩张持续的压力下无以显现。种群虽然也保持膨胀态势,可是明显受制于岳桦的挤压,尤其在海拔较低,气温相对较高,环境更适宜的地方。
图 9 3个高温期前后正常形态长白落叶松和岳桦径级结构趋势线斜率的变化
Fig. 9 The trendline slope change of diameter-class structure of Larix olgensis (general tree) and Betula ermanii on pre- and post-three megathermal periods
1) 长白落叶松垫状生长形海拔最高,其次为旗型树,正常形态海拔最低。而三者所在的海拔范围有着明显的差异,环境梯度差是长白落叶松形态变化的主要因素。长白落叶松通过调整自己的生长方式来实现对不同环境的适应,扩展生存范围。
2) 利用径向与树高生长速度推断长白落叶松与岳桦的竞争力对比结果是长白落叶松以正常生长形态时有较大的竞争力。而随着旗形树的出现,长白落叶松和岳桦的竞争力相似。当长白落叶松变成垫状后,长白落叶松的竞争力应进一步下降,岳桦占优。
3) 长白落叶松和岳桦种群扩张态势的对比结果是随着气温的升高两者种群都膨胀,由于无论何种形态的长白落叶松在幼苗幼树阶段的竞争力不如岳桦,所以总体上长白落叶松种群扩张速度不如岳桦,而且随着气温升温幅度的增加,这种差异也扩大。
4) 虽然长白落叶松在理论上拥有超越岳桦的能力,但实际上很难取代岳桦,所以长白落叶松和岳桦对气候适应表现为林线上侵时以岳桦种群为主力,长白落叶松虽然得到一定程度的扩张,但是由于在初期被岳桦压制很难在林线处大范围展开形成连片的长白落叶松林。
5) 因为长白落叶松以旗形或正常形态生长时竞争力大于岳桦,只要度过幼苗幼树期,岳桦就不可能将其排挤掉,相反长白落叶松的生长优势得以发挥,反过来要限制岳桦的生长,尤其是紧挨大径级长白落叶松的岳桦,因此长白落叶松仍为林线岳桦主要的伴生树种。
1) 长白山自然植被过渡带的性质取决于主要物种的生物学特性和环境因子[2]。本文从径向与树高生长的生物学特征对两种林线乔木种群生长的空间分化进行探讨。长白落叶松因为较强的耐受性[2]成为高纬度广泛分布的林线优势种[1],所以在长白山与林线岳桦优势种形成一定的生态位重叠。另外长白落叶松与岳桦都是阳性树种[2],所以会因争夺空间产生激烈竞争,最后造成生态位的分化而出现生存空间分异[34]。按照生物特征的影响,在林线处最终会形成两种树种在特定空间范围内占有优势而又彼此混杂的植被分布格局。
2) 在生长初期,尤其是幼苗繁殖存活的阶段,岳桦竞争力大于长白落叶松,因此只有岳桦出现较大林窗,岳桦幼苗密度不大,给长白落叶松的幼苗留以充足空间时才能出现长白落叶松挤占岳桦生态位的现象。在岳桦郁闭林中,如果没有大规模的干扰,长白落叶松很难彻底取代岳桦,出现一条完整的林带,这与其他学者的研究结论类似[35]。在长白山建设与开发中,要充分保护岳桦林,如果出现大规模的破坏,则很可能出现岳桦林大片消失的不利情况。
3) 林线动态不仅体现在空间移动的位置变化上,也表现在林线各个组分的变动差异上,各个组分的竞争力强弱不同,对环境变化的反应也有明显差异,这种差异在变化过程中又不是固定不变的,所以在分析气候变化对生态系统影响时不能仅仅观测其空间变化,更应该看到原有系统内部各个组分的变化差异,特别是这种响应差异既受组分本身特点控制也与其他组分的对比有关,还和环境变化幅度与速度都有关系,不但在整体上认识环境变化对生态系统的影响,也应该从内部成分演变的过程中深入剖析对环境变化的响应。
4) 由于受文章篇幅限制,本文只分析林线岳桦和长白落叶松生长的对比关系,没有涉及到所有乔木。同时,对于长白落叶松形态变化的生理机制分析也不够深入,没有从生态因子的对比分析中对这种变化的驱动力进行探讨。今后在研究中还应收集生态因子与长白落叶松、岳桦的生理生态数据,利用一些生态模型从机理上分析两种林线乔木树种的演替规律。
The authors have declared that no competing interests exist.
| [1] |
长白山北坡植物群落ß多样性分析 [J]. |
| [2] |
长白山岳桦种群过渡带位置的研究 [J]. |
| [3] |
长白山森林生态系统服务功能 [J]. |
| [4] |
长白山自然保护区旅游资源开发的生态环境影响及其保护 [J]. |
| [5] |
长白山区域旅游资源开发的几个问题 [J]. |
| [6] |
|
| [7] |
中国东北亚高山林线对全球气候变化的响应 [D]. |
| [8] |
亚高山林线生态交错带的植被生态学研究 [D]. |
| [9] |
长白山北坡落叶松年轮年表及其与气候变化的关系 [J]. |
| [10] |
长白山北坡岳桦种群结构及动态分析 [J]. |
| [11] |
长白山北坡岳桦林线变动的水热条件分析 [J]. |
| [12] |
长白山北坡亚高山云冷杉林的植物种类组成及重要值 [J]. |
| [13] |
气候变化对植物及植被分布的影响研究进展 [J]. |
| [14] |
Growth and structure development of semi-natural larch-spruce-fir (Larix olgensis-Picea jezoensis-Abies nephrolepis) forests in northeast China: 12-year results after thinning [J]. |
| [15] |
气候变暖对长白山主要树种的潜在影响 [J]. |
| [16] |
Simulating carbon balance at treeline for krummholz and dwarf tree growth forms [J]. |
| [17] |
Mapping, modeling, and visualization of the influences of geomorphic processes on the alpine treeline ecotone,Glacier National Park, MT, USA [J].
|
| [18] |
Geomorphological limits to self-organization of alpine forest-tundra ecotone vegetation [J]. |
| [19] |
长白山森林植被NPP主要影响因子的敏感性分析 [J]. |
| [20] |
长白山林线交错带形状与木本植物向苔原侵展和林线动态的关系 [J]. |
| [21] |
长白山北坡植物群落组成、结构及物种多样性的垂直分布 [J]. |
| [22] |
过渡带中岳桦种群生态特征的研究 [J]. |
| [23] |
长白山树线交错带的生物量分配和净生产力 [J]. |
| [24] |
长白山北坡林线岳桦种群与土壤关系的研究 [J]. |
| [25] |
Climatic factors affecting radial growth of Betula ermanii and Betula platypylla in Kamchatka [J]. |
| [26] |
Dendroclimatic analysis of Betula ermanii forests at their upper limit of distribution in Changbai Mountain, Northeast China [J]. |
| [27] |
Comper-assisted quality control in tree-ring dating and measurement [J]. |
| [28] |
东北地带性植被建群种及常见种的分布与水热条件关系的初步研究 [J]. |
| [29] |
长白落叶松林分断面积生长模型的研究 [J]. |
| [30] |
长白山森林植被的生态气候学指标 [J]. |
| [31] |
Relationships of climate change and tree ring of Betula ermanii tree line forest in Changbai Mountain [J]. |
| [32] |
广西南部地区坡地桉树生长与气象因子的关系 [J]. |
| [33] |
Responses of treeislands to air temperature change at treeline on north-facing slopes of the Changbai Mountains [J]. |
| [34] |
长白山北坡木本植物分布与环境关系的典范对应分析 [J]. |
| [35] |
利用影像判读与群落监测分析长白山针叶林动态 [J]. |

/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}