
长白山地土壤甲螨和跳虫多样性的海拔梯度格局
|
刘丹丹 (1993—),女,吉林长春人,博士研究生,主要从事土壤生态学研究。E-mail: liudandan@iga.ac.cn |
收稿日期: 2022-03-12
修回日期: 2022-06-20
网络出版日期: 2023-07-20
基金资助
国家自然科学基金项目(U20A2083)
国家重点研发计划项目(2022YFF1300900)
吉林省科技发展计划项目(20210509037RQ)
版权
Distribution pattern of soil Oribatida and Collembola diversity along altitudinal gradient in the Changbai Mountains
Received date: 2022-03-12
Revised date: 2022-06-20
Online published: 2023-07-20
Supported by
National Natural Science Foundation of China(U20A2083)
National Key R&D Program of China(2022YFF1300900)
Science and Technology Development Program of Jilin Province(20210509037RQ)
Copyright
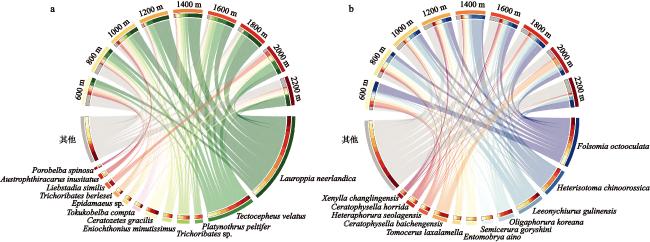
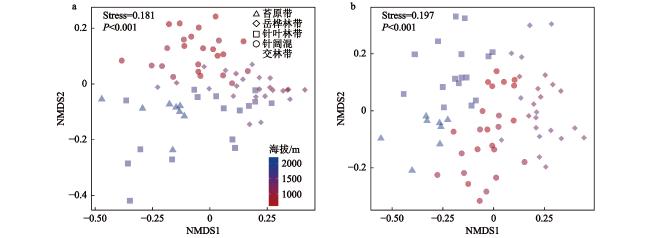
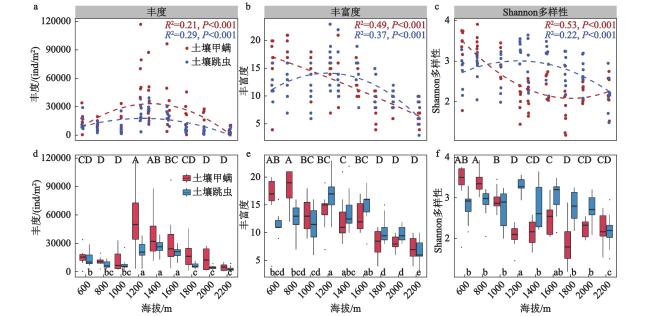
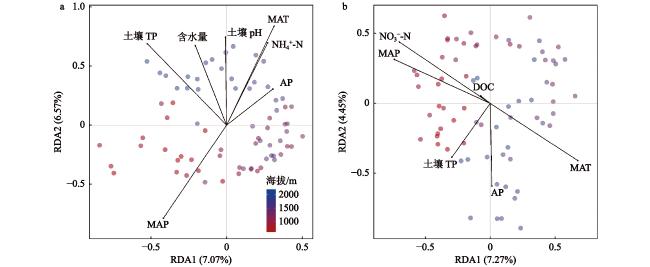
本文围绕物种组成特征、海拔分布格局及其主要驱动因子,探究了长白山北坡9个海拔梯度(600 m、800 m、1000 m、1200 m、1400 m、1600 m、1800 m、2000 m和2200 m)的土壤甲螨和跳虫的多样性分布模式。于2021年7月,共采集到土壤甲螨10688只,隶属于36科45属55种;跳虫6155只,隶属于12科32属54种。非度量多维标度排序(Non-matric multidimentional scaling, NMDS)和多元方差分析表明,土壤甲螨和跳虫的群落结构在9个海拔梯度上存在显著差异,并在长白山4个典型植被带(针阔混交林、针叶林、岳桦林和苔原带)间表现出明显的分离。土壤甲螨和跳虫物种丰度的海拔分布格局呈单峰型,分别在海拔1200 m和1400 m处达到峰值;甲螨的物种丰富度随着海拔升高呈递减分布模式,Shannon多样性指数呈U型分布;而跳虫的丰富度和Shannon多样性指数随海拔升高呈单峰型模式,在海拔1200 m处达到最大值。RDA分析(Redundancy analysis)和VPA分析(Variation partitioning analysis)结果表明,土壤全氮、土壤速效磷以及年均温和年降水量是驱动土壤甲螨和跳虫群落组成沿海拔变化的主要因子。
刘丹丹 , 武海涛 , 于洪贤 , 孙新 , 刘冬 , 程鹏 , 白雪莹 , 戴冠华 , 张仲胜 , 王文锋 . 长白山地土壤甲螨和跳虫多样性的海拔梯度格局[J]. 地理科学, 2023 , 43(7) : 1299 -1309 . DOI: 10.13249/j.cnki.sgs.2023.07.017
The elevational distribution pattern of biodiversity is an essential topic in biogeographic research. As typical representatives of soil mesofauna, Oribatida and Collembola are key biological drivers that regulate soil ecological processes and functions. However, the changes in the vertical zone pattern of soil mesofauna in the Changbai Mountains under climate warming are still poorly understood. This research investigates the biodiversity aggregation patterns of soil oribatid mites and collembola at 9 altitudinal gradients (600 m, 800 m, 1000 m, 1200 m, 1400 m, 1600 m, 1800 m, 2000 m, and 2200 m) on the northern slopes of the Changbai Mountain. In July 2021, a total of 10 688 soil oribatid mites, belonging to 45 genera and 55 species in 36 families, and 6 155 Collembola, belonging to 54 species and 32 genera in 12 families, were collected. NMDS and PERMANOVA showed that the community structure of soil oribatid mites and Collembola differed significantly across the 9 altitudinal gradients and exhibited significant separation between the 4 typical vegetation zones (coniferous and broadleaved forests, mixed coniferous forests, birch forests, and alpine tundra) in the Changbai Mountains. The species richness of oribatid mites exhibited a decreasing distribution pattern with increasing altitude, and the Shannon diversity index showed a U-shaped distribution, while the species richness and the Shannon diversity index of Collembola suggested a hump-shaped with increasing altitude, reaching a maximum at 1200 m above sea level. The results of the RDA and VPA analyses indicated that total soil nitrogen, available phosphorus, mean annual temperature and mean annual precipitation were important drivers that jointly influence the soil mesofauna. Understanding the distribution patterns of soil fauna under changes in elevational gradients can provide a basis for biodiversity conservation and ecological stability studies in forest ecosystems.
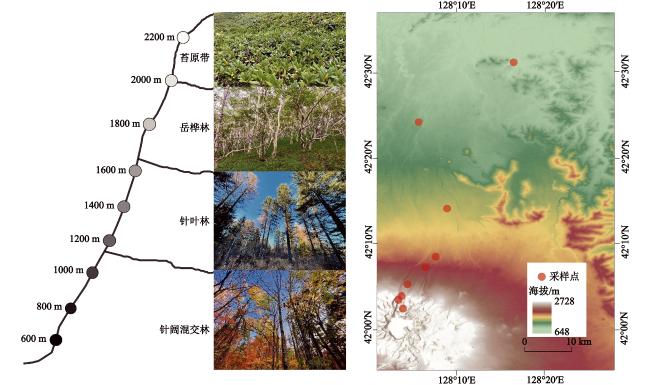
图1 长白山北坡9个海拔梯度采样点分布Fig. 1 Distribution of sampling sites along 9 elevational gradient in the Changbai Mountains |
表1 长白山典型植被带采样点信息Table 1 Summary of the main characteristics of sampling sites in the Changbai Mountains |
| 植被带 | 海拔/m | 经度(E) | 纬度(N) | 年均温/℃ | 年降水量/mm | 优势树种 |
| 针阔混交林 | 600 | 128°16'40″ | 42°31'05″ | 3.15 | 655 | 红松(Pinus koraiensis)、蒙古栎(Quercus mongolica)、水曲柳(Fraxinus mandshurica)、胡桃楸(Juglans mandshurica) |
| 800 | 128°05'46″ | 42°24'08″ | 2.81 | 688 | ||
| 1000 | 128°09'01″ | 42°14'04″ | 1.51 | 708 | ||
| 针叶林 | 1200 | 128°07'42″ | 42°08'27″ | 0.55 | 745 | 长白鱼鳞松(Picea jezoensis)、臭松(Abies nephrolepis)、红松(Pinus koraiensis)、落叶松(Larix olgensis)、蒙古栎(Quercus mongolica)、胡桃楸(Juglans mandshurica) |
| 1400 | 128°06'34″ | 42°07'14″ | 0.05 | 760 | ||
| 1600 | 128°04'29″ | 42°05'15″ | −1.07 | 799 | ||
| 岳桦林 | 1800 | 128°03'49″ | 42°03'53″ | −2.30 | 873 | 岳桦(Betula ermanii)、花楸(Sorbus pohuashanensis)、鱼鳞松(Picea jezoensis)、落叶松(Larix olgensis) |
| 2000 | 128°03'26″ | 42°03'26″ | −3.07 | 913 | ||
| 苔原带 | 2200 | 128°03'57″ | 42°02'25″ | −4.17 | 955 | 高山笃斯(Vaccinium uliginosum)、仙女木(Dryas octopetala L.)、牛皮杜鹃(Rhododendron aureum)、高岭风毛菊(Saussurea tomentosa) |
表2 长白山9个海拔梯度土壤环境因子值(均值±标准差)Table 2 Mean values of soil physical and chemical properties for 9 altitude gradients in the Changbai Mountains |
| 环境因子 | 海拔/m | |||||||||
| 600 | 800 | 1000 | 1200 | 1400 | 1600 | 1800 | 2 000 | 2200 | ||
| 注:单因素方差分析,上标为1;非参数检验,上标为2;***, P < 0.001。 | ||||||||||
| 总碳/ (g/kg)***1 | 57.28±6.88 | 52.03±14.05 | 51.49±14.75 | 41.13±9.17 | 43.58±8.72 | 61.48±14.79 | 55.86±8.26 | 69.70±8.82 | 64.47±9.92 | |
| 可溶性有机碳/ (mg/kg)***2 | 135.11±9.68 | 156.76±34.46 | 246.69±13.08 | 148.98±17.94 | 188.32±29.51 | 206.04±15.37 | 180.68±21.53 | 185.48±39.11 | 155.00±14.50 | |
| 总氮/ (g/kg)***2 | 4.33±0.72 | 4.53±1.32 | 3.75±0.91 | 2.15±0.45 | 2.27±0.37 | 3.47±0.53 | 3.78±0.51 | 4.22±0.96 | 3.92±0.42 | |
| 硝态氮/ (mg/kg)***2 | 9.01±2.90 | 10.74±3.00 | 8.37±0.68 | 10.41±2.81 | 6.37±1.21 | 14.73±2.62 | 17.78±0.85 | 16.76±3.02 | 10.80±2.00 | |
| 铵态氮/ (mg/kg)***2 | 5.19±1.91 | 14.12±4.62 | 7.72±2.51 | 2.62±1.72 | 5.81±1.80 | 9.03±2.26 | 3.18±0.51 | 2.56±1.01 | 1.84±0.66 | |
| 总磷/ (mg/kg)***2 | 1236.18±150.45 | 1086.47±258.33 | 736.08±119.87 | 456.55±80.12 | 470.75±24.19 | 808.15±109.60 | 728.99±61.52 | 789.85±95.72 | 687.46±80.22 | |
| 有效磷/ (mg/kg)***1 | 3.59±0.70 | 13.62±3.08 | 11.30±2.31 | 6.77±2.12 | 6.93±2.35 | 10.23±1.58 | 4.36±1.19 | 3.73±1.97 | 11.93±3.45 | |
| 土壤 pH***2 | 5.92±0.41 | 5.29±0.24 | 5.83±0.13 | 4.99±0.15 | 5.04±0.14 | 5.13±0.20 | 4.73±0.18 | 4.85±0.07 | 5.19±0.12 | |
| 含水量***2 | 61.40±7.72 | 65.65±13.06 | 77.04±10.75 | 36.46±3.74 | 29.19±2.20 | 59.85±7.16 | 47.62±3.75 | 59.36±4.52 | 46.28±6.14 | |
图3 长白山北坡典型植被带土壤甲螨(a)和跳虫(b)群落的非度量多维尺度标序图(NMDS)Fig. 3 Nonmetric multidimensional scaling (NMDS) ordination of Oribatida (a) and Collembola (b) across 4 typical vegetation zones in the Changbai Mountains |
表3 长白山9个海拔梯度土壤甲螨和跳虫的多元方差分析Table 3 Dissimilarity in Oribatida and Collembola community between 9 altitudinal gradients as determined by permutational multivariate analysis of variance |
| 海拔/m | 多元方差分析 | |||||||
| 600 m | 800 m | 1000 m | 1200 m | 1400 m | 1600 m | 1800 m | 2000 m | |
| 注:***表示P<0.001;**表示P<0.01;*表示P<0.05;ns, 表示不显著。 | ||||||||
| 甲螨 Oribatida | ||||||||
| 800 | 0.195*** | |||||||
| 1000 | 0.182** | 0.138* | ||||||
| 1200 | 0.302*** | 0.370** | 0.321** | |||||
| 1400 | 0.313*** | 0.343*** | 0.275** | 0.142ns | ||||
| 1600 | 0.263*** | 0.264*** | 0.217** | 0.150* | 0.172* | |||
| 1800 | 0.225*** | 0.257*** | 0.212** | 0.157* | 0.133ns | 0.145* | ||
| 2000 | 0.217*** | 0.275*** | 0.259*** | 0.289*** | 0.291*** | 0.225** | 0.194** | |
| 2200 | 0.220** | 0.267*** | 0.228*** | 0.374*** | 0.351*** | 0.304*** | 0.241*** | 0.252*** |
| 跳虫 Collembola | ||||||||
| 800 | 0.294** | |||||||
| 1000 | 0.184** | 0.097ns | ||||||
| 1200 | 0.250** | 0.394** | 0.281*** | |||||
| 1400 | 0.380*** | 0.408*** | 0.319*** | 0.279*** | ||||
| 1600 | 0.227*** | 0.327*** | 0.241*** | 0.213*** | 0.289*** | |||
| 1800 | 0.257*** | 0.285*** | 0.199*** | 0.342*** | 0.406*** | 0.235*** | ||
| 2000 | 0.297*** | 0.241*** | 0.196*** | 0.345*** | 0.382*** | 0.275*** | 0.129* | |
| 2200 | 0.291*** | 0.248*** | 0.167** | 0.358*** | 0.403*** | 0.292** | 0.171*** | 0.167** |
| 植被带 | 针阔混交林 | 针叶林 | 岳桦林 | |||||
| 甲螨 Oribatida | ||||||||
| 针叶林 | 0.187*** | |||||||
| 岳桦林 | 0.127*** | 0.108*** | ||||||
| 苔原带 | 0.138*** | 0.246*** | 0.168*** | |||||
| 跳虫 Collembola | ||||||||
| 针叶林 | 0.16*** | |||||||
| 岳桦林 | 0.143*** | 0.211*** | ||||||
| 苔原带 | 0.139*** | 0.223*** | 0.123*** | |||||
表4 长白山土壤甲螨和跳虫环境解释变量的前向选择和变差分解Table 4 Environmental explanatory variables selected by the forward selection process and variation partitioning in the Changbai Mountains |
| 解释部分 | df | Adj.R2 | F | P | 前向选择 | |
| 注:[A],纯土壤环境变量解释量;[B],纯气候变量解释量;[A+B],纯土壤环境变量和纯气候变量的共同解释量;1−[A+B],未解释部分;−为无此项;MAT为年均温、MAP为年降水量。 | ||||||
| 土壤甲螨 | [A] | 5 | 0.123 | 4.21 | 0.001 | 含水量, NH4+-N, TP, AP, pH |
| [B] | 2 | 0.073 | 6.49 | 0.001 | MAT, MAP | |
| [A+B] | 0 | 0.061 | 3.29 | 0.001 | 含水量, NH4+-N, TP, AP, pH, MAT, MAP | |
| 1−[A+B] | − | 0.743 | − | − | − | |
| 土壤跳虫 | [A] | 4 | 0.100 | 3.54 | 0.001 | DOC, NO3−-N, TP, AP |
| [B] | 2 | 0.090 | 5.62 | 0.003 | MAT, MAP | |
| [A+B] | 0 | 0.025 | 3.20 | 0.001 | DOC, NO3−-N, TP, AP, MAT, MAP | |
| 1−[A+B] | − | 0.785 | − | − | − | |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
孙新, 李琪, 姚海凤, 等. 土壤动物与土壤健康[J]. 土壤学报, 2021, 58(5): 1073-1083.
|
| [16] |
|
| [17] |
唐志尧, 方精云. 植物物种多样性的垂直分布格局[J]. 生物多样性, 2004, 12(1): 20-28.
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
谢致敬, 常亮,
|
| [24] |
|
| [25] |
赵淑清, 方精云, 宗占江, 等. 长白山北坡植物群落组成、结构及物种多样性的垂直分布[J]. 生物多样性, 2004, 12(1): 164-173.
|
| [26] |
|
| [27] |
尹文英. 中国土壤动物检索图鉴[M]. 北京: 科学出版社, 1998
|
| [28] |
|
| [29] |
|
| [30] |
刘丹丹, 武海涛, 芦康乐, 等. 空间和环境因子对黄河口自然和淡水恢复湿地底栖动物群落的差异影响[J]. 生态学报, 2021, 41(17): 6893-6903.
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
曹琳琳, 苏宝玲, 周旺明, 等. 长白山区典型植被类型物种多样性空间分布的尺度效应[J]. 生态学杂志, 2017, 36(11): 3102-3108.
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
谷晓楠, 贺红士, 陶岩, 等. 长白山土壤微生物群落结构及酶活性随海拔的分布特征与影响因子[J]. 生态学报, 2017, 37(24): 8374-8384.
|
| [47] |
寇新昌, 殷秀琴. 长白山地不同次生林土壤动物群落多样性特征及其分布格局[J]. 山地学报, 2017, 35(4): 429-436.
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}