
东北长白山泥炭地硅藻−水位埋深转换函数
|
徐双余(2000—),男,云南曲靖人,硕士研究生,主要从事泥炭地硅藻古生态学研究。E-mail: shuangyuxu@cug.edu.cn |
收稿日期: 2023-01-02
修回日期: 2023-04-11
网络出版日期: 2024-08-21
基金资助
国家自然科学基金项目(U20A2094)
国家自然科学基金项目(42171166)
国家自然科学基金项目(41572343)
版权
Diatom-based water-table depth transfer function from peatlands in the Changbai Mountains, Northeast China
Received date: 2023-01-02
Revised date: 2023-04-11
Online published: 2024-08-21
Supported by
National Natural Science Foundation of China(U20A2094)
National Natural Science Foundation of China(42171166)
National Natural Science Foundation of China(41572343)
Copyright
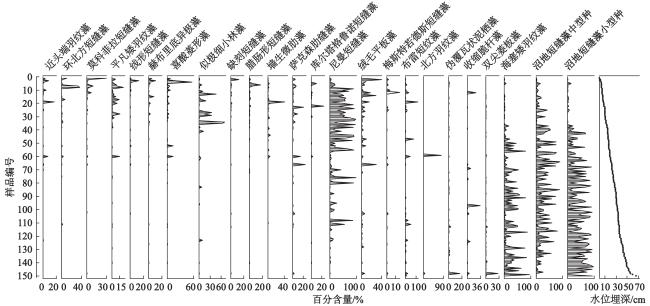
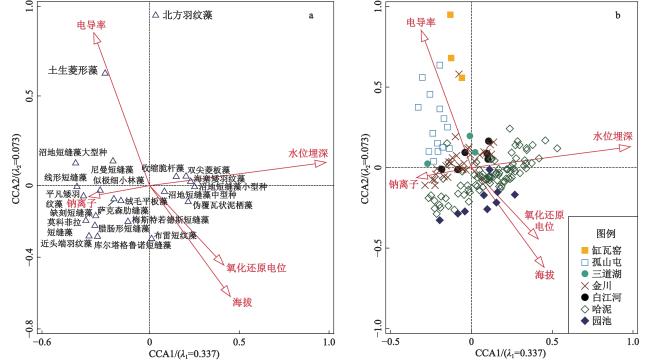
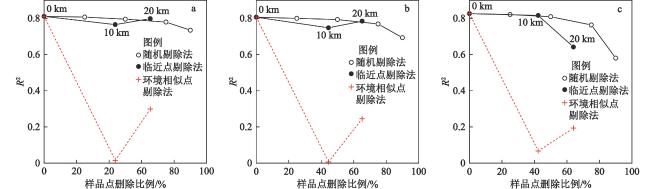
本研究基于2016、2019和2021年夏季在东北长白山地区7处泥炭地采集的160个表层样品和环境因子数据,结合表层样品硅藻分析,构建了硅藻属种与环境因子数据库,该数据库包括64个硅藻属种和11个环境变量。优势硅藻属种组合沿水位埋深梯度(0~66 cm)呈现明显分异,偏典范对应分析结果表明,水位埋深单独解释了硅藻组合数据总方差的8.4%,远大于其他环境因子的单独解释份额。基于此,选取加权平均模型和最大似然估计模型构建硅藻−水位转换函数。不同模型分析结果显示,典型回归耐受值降权加权平均模型构建的硅藻−水位转换函数表现最佳,剔除残差大于20%水位梯度(13.2 cm)的样品后进行留一法交叉检验,该模型的推导值与实测值相关系数最大和推导误差最小。本文构建的硅藻−水位转换函数为定量反演长白山地区泥炭地过去水位变化提供了新的有效方法。
徐双余 , 卜兆君 , 黄咸雨 , 彭佳 , 李俊璐 , 陈旭 . 东北长白山泥炭地硅藻−水位埋深转换函数[J]. 地理科学, 2024 , 44(8) : 1459 -1469 . DOI: 10.13249/j.cnki.sgs.20221516
Peatlands, as unique limno-terrestrial ecosystems, play an important role in water storage, biodiversity conservation and global carbon cycling. Water table is a key environmental variable controlling the biogeochemical process in peatlands, and hence it is of great significance to reveal water-table changes for further understanding the evolution of peatlands. Based on one hundred and sixty samples collected from seven peatlands in the Changbai Mountains, Northeast China, in the summer of 2016, 2019 and 2021, this study established a dataset consisting of sixty-four diatom species and eleven environmental variables. The diatom assemblages registered clear variations along the water-table gradient from 0 to 66 cm. Partial CCA revealed that Depth to the Water Table (DWT) independently explained 8.4% of the total variance in diatom composition, which was much higher than the sole effect of other environmental factors. Diatom-based water-table transfer function was developed using the Weighted Averaging model and the Maximum Likelihood model, respectively. Our results showed that tolerance downweighted Weighted Averaging model with classical deshrinking (WA.cla.tol) had the best performance. After removing the samples with residuals >20% of total measured water table range (13.2 cm), the WA.cla.tol model yielded the highest coefficient of determination (R2) between diatom-inferred DWT and observed DWT, and the lowest Root Mean Square Error of Prediction (RMSEP) in leave-one-out cross validation. The diatom-based water-table transfer function built in this study provides a new and effective method for quantitative reconstruction of water-table changes in peatlands of the Changbai Mountains.
表1 显著环境因子的边缘效应和独立效应Table 1 Marginal effects and unique effects of each significant environmental variable |
| 环境因子 | 边缘效应 | 独立效应 | ||||
| 特征值λ1 | 特征值λ2 | λ1/λ2 | 总方差解释份额/% | P值 | ||
| 注:DWT为水位埋深;Alt为海拔;Cond为电导率;Na+为钠离子含量;ORP为氧化还原电位;λ1、λ2分别为第一特征值、第二特征值。 | ||||||
| DWT | 0.327 | 0.294 | 1.11 | 8.4 | 0.001 | |
| Alt | 0.122 | 0.495 | 0.25 | 2.0 | 0.001 | |
| Cond | 0.094 | 0.510 | 0.18 | 1.6 | 0.001 | |
| Na+ | 0.065 | 0.498 | 0.13 | 0.6 | 0.015 | |
| ORP | 0.088 | 0.482 | 0.18 | 0.6 | 0.025 | |
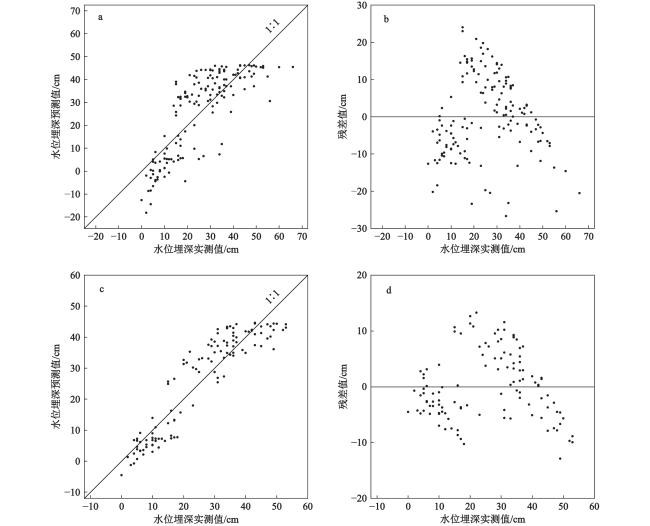
表2 硅藻−水位转换函数的推导能力评估结果Table 2 The performance statistics for the diatom-based water table transfer function |
| 优化前模型 | RMSEP | R2 | 最大推导误差 |
| 注:括号外的数字为LOO验证结果;括号内数字为LGO验证结果;n为剔除的样品数;RMSEP为预测均方根误差。 | |||
| WA.cla.tol | 11.30 (11.48) | 0.60 (0.49) | 19.70 (28.15) |
| WA.inv.tol | 9.32 (11.53) | 0.59 (0.41) | 25.97 (34.93) |
| WA.inv | 9.49 (11.63) | 0.58 (0.40) | 26.47 (34.53) |
| WA.cla | 11.68 (11.71) | 0.58 (0.49) | 20.33 (26.84) |
| ML | 11.66 (11.53) | 0.56 (0.46) | 19.70 (27.73) |
| 优化后模型 | RMSEP | R2 | 最大推导误差 |
| WA.cla.tol (n=34) | 6.34 (7.31) | 0.83 (0.78) | 8.70 (14.46) |
| ML (n=30) | 6.60 (8.45) | 0.83 (0.70) | 10.32 (36.07) |
| WA.cla (n=40) | 6.88 (8.20) | 0.81 (0.71) | 10.06 (15.69) |
| WA.inv (n=22) | 6.56 (7.78) | 0.76 (0.69) | 12.33 (17.61) |
| WA.inv.tol (n=18) | 6.71 (8.14) | 0.75 (0.65) | 12.55 (18.30) |
| [1] |
Rydin H, Jeglum J K. The Biology of Peatlands[M]. Oxford: Oxford University Press, 2013.
|
| [2] |
Minasny B, Berglund Ö, Connolly J et al. Digital mapping of peatlands—A critical review[J]. Earth-Science Reviews, 2019, 196: 102870.
|
| [3] |
Waddington J M, Morris P J, Kettridge N et al. Hydrological feedbacks in northern peatlands[J]. Ecohydrology, 2015, 8(1): 113-127.
|
| [4] |
Chen X, McGowan S, Bu Z J et al. Diatom-inferred microtopography formation in peatlands[J]. Earth Surface Processes and Landforms, 2022, 47(2): 672-687.
|
| [5] |
Loisel J, Gallego-Sala A. Ecological resilience of restored peatlands to climate change[J]. Communications Earth & Environment, 2022, 3(1): 208.
|
| [6] |
Fenner N, Freeman C. Drought-induced carbon loss in peatlands[J]. Nature Geoscience, 2011, 4(12): 895-900.
|
| [7] |
Chen X, McGowan S, Bu Z et al. Diatom-based water-table reconstruction in Sphagnum peatlands of Northeastern China[J]. Water Research, 2020, 174: 115648.
|
| [8] |
Chambers F M, Booth R K, De Vleeschouwer F et al. Development and refinement of proxy-climate indicators from peats[J]. Quaternary International, 2012, 268: 21-33.
|
| [9] |
Blundell A, Barber K. A 2800-year palaeoclimatic record from Tore Hill Moss, Strathspey, Scotland: The need for a multi-proxy approach to peat-based climate reconstructions[J]. Quaternary Science Reviews, 2005, 24(10-11): 1261-1277.
|
| [10] |
Mitchell E, Payne R J, van der Knaap W O et al. The performance of single- and multi-proxy transfer functions (testate amoebae, bryophytes, vascular plants) for reconstructing mire surface wetness and pH[J]. Quaternary Research, 2013, 79(1): 6-13.
|
| [11] |
Yang Q, Li H, Zhao H et al. Plant assemblages-based quantitative reconstruction of past mire surface wetness: A case study in the Changbai Mountains region, Northeast China[J]. Catena, 2022, 216: 106412
|
| [12] |
Liu H, Gu Y, Lun Z et al. Phytolith-inferred transfer function for paleohydrological reconstruction of Dajiuhu peatland, central China[J]. The Holocene, 2018, 28(10): 1623-1630.
|
| [13] |
Qin Y, Li H, Mazei Y et al. Developing a continental-scale testate amoeba hydrological transfer function for Asian peatlands[J]. Quaternary Science Reviews, 2021, 258: 106868
|
| [14] |
Huang X, Pancost R D, Xue J et al. Response of carbon cycle to drier conditions in the mid-Holocene in central China[J]. Nature Communications, 2018, 9(1): 1369.
|
| [15] |
Rioual P, Lu Y, Yang H et al. Diatom-environment relationships and a transfer function for conductivity in lakes of the Badain Jaran Desert, Inner Mongolia, China[J]. Journal of Paleolimnology, 2013, 50(2): 207-229.
|
| [16] |
Yang X, Anderson N J, Dong X et al. Surface sediment diatom assemblages and epilimnetic total phosphorus in large, shallow lakes of the Yangtze floodplain: Their relationships and implications for assessing long-term eutrophication[J]. Freshwater Biology, 2008, 53(7): 1273-1290.
|
| [17] |
陈小林, 陈光杰, 刘园园, 等. 云南45个湖泊硅藻−总磷转换函数及其定量重建评价[J]. 湖泊科学, 2022, 35(1): 1-18.
Chen Xiaolin, Chen Guangjie, Liu Yuanyuan et al. Evaluation of the quantitative relationships between diatom communities and total phosphorus (TP) in 45 lakes and their applications for TP reconstruction in Yunnan, Southwest China. Journal of Lake Sciences, 2022, 35(1): 1-18.
|
| [18] |
Gaiser E, Rühland K. Diatoms as indicators of environmental change in wetlands and peatlands[M]// Smol J P et al. The diatoms: Applications for the environmental and earth sciences. Cambridge: Cambridge University Press, 2010: 473-493.
|
| [19] |
Poulíčková A, Hajkova P, Krenkova P et al. Distribution of diatoms and bryophytes on linear transects through spring fens[J]. Nova Hedwigia, 2004, 78(3-4): 411-424.
|
| [20] |
Chen X, Bu Z, Yang X et al. Epiphytic diatoms and their relation to moisture and moss composition in two montane mires, Northeast China[J]. Fundamental and Applied Limnology, 2012, 181(3): 197-206.
|
| [21] |
Hargan K E, Ruhland K M, Paterson A M et al. The influence of water-table depth and pH on the spatial distribution of diatom species in peatlands of the Boreal Shield and Hudson Plains, Canada[J]. Botany, 2015, 93(2): 57-74.
|
| [22] |
Ma L, Gao C, Kattel G R et al. Evidence of Holocene water level changes inferred from diatoms and the evolution of the Honghe Peatland on the Sanjiang Plain of Northeast China[J]. Quaternary International, 2018, 476: 82-94.
|
| [23] |
Payne R J, Telford R J, Blackford J J et al. Testing peatland testate amoeba transfer functions: Appropriate methods for clustered training-sets[J]. Holocene, 2012, 22(7): 819-825.
|
| [24] |
Telford R J, Birks H. Effect of uneven sampling along an environmental gradient on transfer-function performance[J]. Journal of Paleolimnology, 2011, 46(1): 99-106.
|
| [25] |
Telford R J, Birks H. Evaluation of transfer functions in spatially structured environments[J]. Quaternary Science Reviews, 2009, 28(13-14): 1309-1316.
|
| [26] |
郎惠卿. 中国湿地植被[M]. 北京: 科学出版社, 1999
Lang Huiqing. Wetland vegetation in China. Beijing: Science Press, 1999.
|
| [27] |
Li H, Wang S, Zhao H et al. A testate amoebae transfer function from Sphagnum-dominated peatlands in the Lesser Khingan Mountains, NE China[J]. Journal of Paleolimnology, 2015, 54(2-3): 189-203.
|
| [28] |
Kulikovskiy M S, Lange-Bertalot H, Witkowski A et al. Diatom assemblages from Sphagnum bogs of the World. I. Nur bog in northern Mongolia[M]. Stuttgart, Germany: J. Cramer, 2010.
|
| [29] |
Lange-Bertalot H, Bak M, Witkowski A et al. Diatoms of Europe. Vol. 6 Eunotia and some related genera[M]. Ruggell: ARG Gantner Verlag KG, 2011.
|
| [30] |
Šmilauer P, Lepš J. Multivariate analysis of ecological data using Canoco 5[M]. Cambridge: Cambridge University Press, 2014.
|
| [31] |
Juggins S. Quantitative reconstructions in palaeolimnology: New paradigm or sick science?[J]. Quaternary Science Reviews, 2013, 64: 20-32.
|
| [32] |
李鸿凯, 李微微, 蒲有宝, 等. 应用rioja软件包建立有壳变形虫−环境因子转换函数[J]. 地理科学, 2013, 33(8): 1022-1028.
Li Hongkai, Li Weiwei, Pu Youbao et al. Building transfer functions between testate amoeba and environmental variables with ‘rioja’ package. Scientia Geographica Sinica, 2013, 33(8): 1022-1028.
|
| [33] |
Juggins S, Birks H J B. Quantitative environmental reconstructions from biological data[M]// Birks H J B, Lotter A F, Juggins S et al . Tracking environmental change using lake rediments: Data handling and numerical techniques. Dordrecht: Springer Netherlands, 2012: 431-494.
|
| [34] |
Juggins S. rioja: Analysis of quaternary science data, R package version (0.9-26)[CP/OL]. 2020. http://cran.r-project.org/package=rioja.
|
| [35] |
Nychka D, Furrer R, Paige J et al. fields: Tools for spatial data. R package version 13.3[CP/OL]. 2021. https://github.com/dnychka/fieldsRPackage.
|
| [36] |
Telford R J. PalaeoSig: Significance tests of quantitative palaeoenvironmental reconstructions, R package version (2.0-3)[CP/OL]. 2019. http://cran.r-project.org/package=palaeoSig.
|
| [37] |
R Core Team. R: A language and environment for statistical computing[CP/OL]. 2021. https://www.R-project.org/.
|
| [38] |
Jirousek M, Poulickova A, Kintrova K et al. Long-term and contemporary environmental conditions as determinants of the species composition of bog organisms[J]. Freshwater Biology, 2013, 58(10): 2196-2207.
|
| [39] |
Kokfelt U, Struyf E, Randsalu L. Diatoms in peat-Dominant producers in a changing environment?[J]. Soil Biology and Biochemistry, 2009, 41(8): 1764-1766.
|
| [40] |
Van de Vijver B, Beyens L. The epiphytic diatom flora of mosses from Strømness Bay area, South Georgia[J]. Polar Biology, 1997, 17(6): 492-501.
|
| [41] |
Buczkó K, Wojtal K. Moss inhabiting siliceous algae from Hungarian peat bogs[J]. Studia Botanica Hungarica, 2005, 36: 21-42.
|
| [42] |
Cantonati M, Lange-Bertalot H, Decet F et al. Diatoms in very-shallow pools of the site of community importance Danta di Cadore Mires (south-eastern Alps), and the potential contribution of these habitats to diatom biodiversity conservation[J]. Nova Hedwigia, 2011, 93(3-4): 475-507.
|
| [43] |
Chen X, Qin Y, Stevenson M A et al. Diatom communities along pH and hydrological gradients in three montane mires, central China[J]. Ecological Indicators, 2014, 45: 123-129.
|
| [44] |
Gremmen N, Van De Vijver B, Frenot Y et al. Distribution of moss-inhabiting diatoms along an altitudinal gradient at sub-Antarctic Iles Kerguelen[J]. Antarctic Science, 2007, 19(1): 17-24.
|
| [45] |
Lotter A F, Birks H, Hofmann W et al. Modern diatom, cladocera, chironomid, and chrysophyte cyst assemblages as quantitative indicators for the reconstruction of past environmental conditions in the Alps. I. Climate[J]. Journal of Paleolimnology, 1997, 18(4): 395-420.
|
| [46] |
Anderson N J. Diatoms, temperature and climatic change[J]. European Journal of Phycology, 2000, 35(4): 307-314.
|
| [47] |
Clymo R S, Hayward P M. The Ecology of Sphagnum[M]// Smith A J E. Bryophyte Ecology. Netherlands: Springer, 1982: 229-289.
|
| [48] |
Fránková M, Bojkova J, Poulickova A et al. The structure and species richness of the diatom assemblages of the Western Carpathian spring fens along the gradient of mineral richness[J]. Fottea, 2009, 9(2): 355-368.
|
| [49] |
Han D, Sun Y, Cong J et al. Ecological distribution of modern diatom in peatlands in the northern Greater Khingan Mountains and its environmental implications[J]. Environmental Science and Pollution Research, 2023, 30: 36607-36618.
|
| [50] |
Charman D J, Blundell A. A new European testate amoebae transfer function for palaeohydrological reconstruction on ombrotrophic peatlands[J]. Journal of Quaternary Science, 2007, 22(3): 209-221.
|
| [51] |
Zhang H, Amesbury M J, Ronkainen T et al. Testate amoeba as palaeohydrological indicators in the permafrost peatlands of north-east European Russia and Finnish Lapland[J]. Journal of Quaternary Science, 2017, 32(7): 976-988.
|
| [52] |
Xu S, Huang B, Zeng L et al. Diatom cell-size composition as a novel tool for quantitative estimates of the water table in peatlands[J]. Biology Letters, 2024(20): 20240062
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}