靳英华 , 许嘉巍, 刘丽娜
, 许嘉巍, 刘丽娜
Jin Yinghua, Xu Jiawei, Liu Lina
中图分类号: Q948
文献标识码: A
文章编号: 1000-0690(2016)08-1212-07
通讯作者:
收稿日期: 2015-10-16
修回日期: 2015-12-23
网络出版日期: 2016-08-20
版权声明: 2016 《地理科学》编辑部 本文是开放获取期刊文献,在以下情况下可以自由使用:学术研究、学术交流、科研教学等,但不允许用于商业目的.
基金资助:
作者简介:
作者简介:靳英华(1968-),女,博士,副教授,主要从事自然地理学研究。E-mail: jinyh796@nenu.edu.cn
展开
摘要
通过开展植被样方调查,确定苔原带的主要优势种,分析优势种的分布格局及其关联性,从地理角度分析长白山苔原带植被变化特征。研究表明:① 长白山苔原带8个优势种中,草本植物已占6个。灌木中牛皮杜鹃(Rhododendron chrysanthum)依然为最主要的优势种,但笃斯越橘(Vaccinium uliginosum)的优势地位已被草本的小白花地榆(Sanguisorba parviflora)取代。草本植物聚集程度强,苔原植被存在草甸化趋势。② 除牛皮杜鹃与大白花地榆(Sanguisorba stipulata)之间存在较显著的负联结,排它现象明显外,多种优势种之间存在正联结,有弱的伴生现象,牛皮杜鹃和小白花地榆为强正联结,伴生现象更明显,草本植物与灌木伴生将长期存在。③ 各草本植物入侵、定植、扩展机制多样。小白花地榆分布广,聚集程度较弱,以种子繁殖为主,扩展速度较快;大白花地榆、小叶章(Calamagrostis angustifolia)等聚集程度较强,以无性繁殖为主,扩展速度较慢,但竞争能力强,在适宜生境中,能驱除其它物种。
关键词:
Abstract
Ecological environment and plant community has changed obviously to tundra zone in western slope of the Changbai Mountains during the last three decades. The space of herbaceous plant extended obviously, but the distribution range of shrub atrophied. In order to explore the tundra vegetation change characteristic from the perspective of geography, we carried out vegetation investigation to reveal the change, spatial distribution pattern and associations of the main dominant species of tundra zone in the Changbai Mountains. The results showed that: 6 dominant plant species are herbaceous plant among the 8 dominant plant species in the tundra zone of the Changbai Mountains. Although Rhododendron chrysanthum is still the main dominant plant species of shrubs and has a higher species abundance, a lower aggregation degree and occupies most habitats, but the dominant position of Vaccinium uliginosum has declined, living space has reduced. Sanguisorba parviflora has a high species-abundance, a low degree of aggregation. Sanguisorba parviflora and Rhododendron chrysanthum was significantly positively associated, and Sanguisorba parviflora became a strong extension of herbaceous plant. Herbaceous plants such as Sanguisorba stipulata and Calamagrostis angustifolia have a high degree of aggregation, present patch distribution, and have occupied part of habitats. The tundra vegetation of the Changbai Mountains has the trend of meadowization. Among mostly dominant plants showed a low correlation and had a higher degree of aggregation. It illustrates that each dominate plant specie has different choice of habitats and the pressure of distribution change get small because of inter-specific competition, so the change of tundra vegetation has stabilized. Except Rhododendron chrysanthum and Sanguisorba stipulata showed a significant negative correlation and had a obviously exclusive phenomenon. Among several dominant plant species showed a positive correlation and had a slight attendant phenomenon. Rhododendron chrysanthum and Sanguisorba parviflora showed a significant positive correlation and attendant phenomenon. It is visible that herbaceous plant and shrubs associated will exist for a long time. The herbaceous plant has a diverse invasion, colonization, and expansible mechanism. Sanguisorba parviflora has wide distribution, a low aggregation degree, because the predominant propagation mode of Sanguisorba parviflora was seed reproduces, it should have a faster extended speed. The predominant propagation mode of Sanguisorba stipulata and Calamagrostis angustifolia was asexual, their extended speed is slower, but inter-specific competition ability was stronger, in adaptation habitats, they can get rid of other species effectively.
Keywords:
过去30 a,长白山苔原带的生态环境已发生了明显的变化,生长季温度显著升高,积温增加,生长期延长;降水量增多,降水日数减少,降水强度增加;积雪厚度下降,融雪期提前,雪被期缩短[1~10]。与此同时,长白山苔原带已经观测到氮沉降量增加,尤其在公路附近最为显著[11]。
将近年的植物样方调查结果与钱家驹和钱宏的记载对比后发现[12,13],长白山西坡苔原带植被发生了显著的变化[14~16],一些草本植物如大白花地榆(Sanguisorba stipulata),小白花地榆(Sanguisorba parviflora),小叶章(Calamagrostis angustifolia),单花橐吾(Ligularia jamesii),高岭风毛菊(Saussurea tomentosa),长白老鹳草(Geranium baishanense)、尖被藜芦(Veratrum oxysepalum)、大苞柴胡(Bupleurum euphorbioides)等多度增加,分布范围显著扩展。这些或由岳桦林带迁入或仅是苔原带的偶见种,现已形成草本植物斑块,成为常见种甚至优势种;与之相反,以灌木为代表的苔原优势种和特有种如牛皮杜鹃(Rhododendron chrysanthum)、笃斯越橘(Vaccinium uliginosum)、松毛翠(Phyllodoce caerulea)、宽叶仙女木(Dryas octopetala)、毛毡杜鹃(Rhododendron confertissimum)等的分布区明显萎缩[17]。
目前关于长白山苔原带植被变化机理尚不清楚,本研究旨在揭示长白山苔原带主要优势种的变化、优势种的空间分布格局及其关联性,从地理角度分析长白山苔原带植被变化特征。
长白山地处吉林省东南(41°23'N~42°36'N,126°55'E~129°E),为中国和朝鲜两国的界山,是中国东北最高山峰(海拔2 691 m)。由于地势高峻,气候、植被和土壤成明显的垂直带分布。从下而上为山地针阔混交林带、山地针叶林带、亚高山岳桦林带和高山苔原带。
长白山苔原带位于海拔2 000 m以上的火山锥上部。地表多为碱性粗面岩风化物和少量火山灰,地貌为流水改造的火山锥坡面,土壤为薄层山地苔原土,寒带气候。研究区植物中极地或高山种属约占80%,以灌木为主,牛皮杜鹃群落和笃斯越橘群落为苔原带最常见的植物群落,群落中存在2个层片,为灌木层片、苔藓-地衣层片,一般不形成草本植物层片。
1.2.1 样地调查
野外调查采用设置样地进行系统抽样的方法。 2014年8月在长白山西坡苔原带布设了100 m×1 600 m的大样地,样地具体位置见表1。在此大样方内,沿样带主方向每隔50 m,横向设置样条,均匀布设4个1 m×1 m的控制性样方,共计33×4=132个控制样方,记录经纬度、海拔(由GPS测得)、坡度和坡向(由罗盘测得),进行植被样方调查(植物种、株高、株数、盖度)。
表1 样地位置
Table 1 Location of sample plots
| 控制点 | 纬度 | 经度 | 海拔(m) |
|---|---|---|---|
| 样带西南角 | 41.9895°N | 128.0031°E | 2073 |
| 样带西北角 | 41.9900°N | 128.0023°E | 2080 |
| 样带东南角 | 41.9931°N | 128.0190°E | 2293 |
| 样带东北角 | 41.9938°N | 128.0187°E | 2322 |
1.2.2 数据处理
1) 优势种的确定。计算长白山苔原带中各物种的重要值,再用降序排列的重要值来确定物种的优势程度,其中,重要值数值大于3的物种判定为优势种。
重要值=[相对密度+相对频度+相对盖度]/3。其中,相对密度=某种植物的个体数/全部植物个体数×100;相对频度=某种植物的种群频度/所有种群频度之和×100;相对盖度=某种植物的种群盖度/所有种群盖度之和×100。
2) 分布格局类型与聚集强度的判定。采用扩散系数(DI) 测定分布格局类型, 然后用t检验判断分布格局的显著性。
式中,S2为种群多度的方差,
采用聚集强度负二项参数(K)判定聚集强度。
式中,负二项参数K值与种群密度无关,K值愈小,聚集度愈大。如果K值趋于无穷大(一般为8以上) ,则接近随机分布。
3) 种间关联研究。在调查样地中选取优势度相对较大的物种进行种间联结及相关性分析。建立2×2联列表(表2),并根据原始资料矩阵,计算各种对的 a、b、c和d值。以λ2检验为基础,结合连接系数AC和共同出现百分率PC等方法共同测定物种的联结性。
① 检验2个种关联与否λ2检验。
表2和(3)式中,a为2个种均出现的样方数,b为仅有A种出现的样方数,c为仅有B种出现的样方数,d为2种都未出现的样方数。式中,N为小样方总数,λ2 近似遵从自由度为1的分布,当λ2<3.84(p>0.05)时,种间联结独立;当3.84≤λ2<6.635(0.01<p<0.05)时,种间有一定的生态联结;当λ2≥6.635(p<0.01)时,种间有显著的生态联结。λ2本身没有负值,判定正、负联结的方法是当ad>bc,种间具正关联;若ad<bc,种间具负关联。
表2 两物种的2×2联列表
Table 2 The 2×2 contingency table of two species
| B种 | |||
|---|---|---|---|
| A种 | 出现 | 不出现 | 合计 |
| 出现 | a | b | a+b |
| 不出现 | c | d | c+d |
| 合计 | a+c | b+d | N=a+b+c+d |
② 联结系数AC。为进一步测定种间关联强度,采用种间联结系数AC,AC用来进一步检验由λ2所测出的结果及说明种间联结程度。AC的值域为[-1,1],AC值越趋近1,种对的正关联性越强,AC值越趋近-1,种对负关联性越强,AC值为0,种间完全独立。其计算公式为
③ 共同出现百分率PC。式(5)用来测度物种间正联结程度的,PC的值域为[0,1],值越趋近于1则表明该种对的正联结越紧密。
1.2.3 统计方法
利用Excel软件对各物种的密度、频度和盖度进行统计,计算各物种的重要值,确定苔原带优势物种;利用SPSS 18.0对苔原优势植物进行聚类分析。
2.1.1 物种组成
长白山苔原带大样地内植物种类共78种,隶属22科。包括杨柳科(Salicaceae)、蓼科(Polygonaceae)、石竹科(Caryophyllaceae)、毛茛科(Ranunculaceae)、景天科(Crassulaceae)、虎耳草科(Saxifragaceae)、蔷薇科(Rosaceae)、豆科(Leguminosae)、牻牛儿苗科(Geraniaceae)、堇菜科(Violaceae)、伞形科(Umbelliferae)、杜鹃花科(Ericaaeae)、报春花科(Primulaceae)、龙胆科(Gentianaceae)、玄参科(Scrophulariaceae)、桔梗科(Campanulaceae)、菊科(Compositae)、禾本科(Gramineae)、莎草科(Cyeraceae)、灯心草科(Juncaceae)、百合科(Liliaceae)、兰科(Orchidaceae)。
2.1.2 主要优势物种
长白山高山苔原带植物群落中有30个种群的重要值超过0.5,选取重要值>3.0的8个种群为苔原带优势种(表3),按重要值大小依次为牛皮杜鹃、大白花地榆、小白花地榆、小叶章、单花橐吾、高岭风毛菊、笃斯越橘、长白老鹳草。
表3 长白山高山苔原带植物群落各种群重要值
Table 3 Each population IV of the alpine tundra vegetation population of the Changbai Mountains
| 序号 | 总株数 | 相对密度(RD) | 相对盖度(RC) | 相对频度(RF) | 重要值(IV) | |
|---|---|---|---|---|---|---|
| 1 | 牛皮杜鹃 | 7028 | 30.12 | 24.08 | 7.48 | 20.56 |
| 2 | 大白花地榆 | 2281 | 9.78 | 11.57 | 5.30 | 8.88 |
| 3 | 小白花地榆 | 2583 | 11.07 | 7.21 | 7.48 | 8.59 |
| 4 | 小叶章 | 1872 | 8.02 | 8.30 | 3.95 | 6.76 |
| 5 | 单花橐吾 | 1080 | 4.63 | 4.83 | 4.62 | 4.70 |
| 6 | 高岭风毛菊 | 747 | 3.20 | 3.18 | 5.04 | 3.81 |
| 7 | 笃斯越橘 | 635 | 2.72 | 5.63 | 2.52 | 3.62 |
| 8 | 长白老鹳草 | 661 | 2.83 | 2.08 | 4.29 | 3.07 |
长白山西坡优势植物的方差/均值比均大于1,且都通过t检验,可以判定8种优势植物的分布格局都属于聚集分布(表4)。牛皮杜鹃的K值最大,聚集强度最弱;笃斯越橘的K值最小,聚集强度最强。除了小白花地榆,大多数草本植物的K值都小,因此聚集强度都较强,聚集强度由弱到强依次为小白花地榆、大白花地榆、高岭风毛菊、长白老鹳草、小叶章和单花橐吾(表4)。
表4 优势植物空间格局类型
Table 4 Spatial pattern type of dominant species
| 植物种名 | DI | T_检验 | K | 格局类型 | 多度 |
|---|---|---|---|---|---|
| 牛皮杜鹃 | 53.85 | 126.44 | 1.02 | 聚集 | 53.24 |
| 大白花地榆 | 43.69 | 104.52 | 0.4 | 聚集 | 17.28 |
| 小白花地榆 | 21.60 | 51.67 | 0.97 | 聚集 | 19.57 |
| 小叶章 | 57.95 | 138.64 | 0.25 | 聚集 | 14.18 |
| 单花橐吾 | 37.23 | 89.07 | 0.23 | 聚集 | 8.18 |
| 高岭风毛菊 | 16.13 | 38.59 | 0.38 | 聚集 | 5.66 |
| 笃斯越橘 | 50.47 | 120.74 | 0.1 | 聚集 | 4.81 |
| 长白老鹳草 | 19.61 | 46.91 | 0.27 | 聚集 | 5 |
虽然目前优势种都呈聚集分布,但是其分布特点还是有差异,通过聚类分析,可分成4种类型:① 多度大,聚集程度弱,如:牛皮杜鹃。② 多度较大,聚集强度较弱,如:小白花地榆。③ 多度较大,聚集强度大,如:小叶章和大白花地榆。④ 多度小,聚集强度大,如:笃斯越橘、高岭风毛菊、长白老鹳草和单花橐吾。
研究区8个优势种组成的28个种对中,联结性达到极显著(λ2>6.635)和显著水平(3.841< λ2< 6.635)的有16对;其中检验成正联结的有16对,占总对数的 57.1%,负联结的有12对,占总对数的42.9%(表5)。
表5 优势种群种间关联的λ2值测定矩阵表
Table 5 The λ2 mensuration matrix of interspecific association for dominant species pairs
| 牛皮杜鹃 | 大白花地榆 | 小白花地榆 | 小叶章 | 单花橐吾 | 高岭风毛菊 | 笃斯越橘 | |
|---|---|---|---|---|---|---|---|
| 大白花地榆 | 27.009 - | ||||||
| 小白花地榆 | 65.362 + | 27.525 - | |||||
| 小叶章 | 1.877 - | 9.765 + | 1.608 - | ||||
| 单花橐吾 | 8.161 + | 25.369 + | 7.040 - | 0.191 + | |||
| 高岭风毛菊 | 19.166 + | 35.965 - | 21.018 + | 14.582 - | 7.071- | ||
| 笃斯越橘 | 5.460+ | 33.015 - | 6.819 + | 0.896 - | 25.557 - | 5.982 + | |
| 长白老鹳草 | 3.804 + | 1.889 - | 8.945 + | 0.015 + | 0.054 + | 13.720 + | 0.002 + |
牛皮杜鹃为苔原带最主要的优势种,与灌木笃斯越橘显著正联结;与6种草本优势种的关联性不同,与小白花地榆、单花橐吾和高岭风毛菊为极显著的正联结,与小叶章、长白老鹳草间联结独立,与大白花地榆极显著的负联结。笃斯越橘与草本优势植物的关联性和牛皮杜鹃与草本的关联性基本一致。
草本优势植物的15个种对,有5对是极显著和显著的正联结,5对是极显著的负联结,5对联结独立(表5)。小叶章、长白老鹳草与大部分草本优势植物联结独立;小白花地榆仅与小叶章联结独立,与其它草本优势种为显著的联结;高岭风毛菊和所有草本优势植物都为极显著的联结;大白花地榆、单花橐吾与其它草本优势植物都有显著的联结。
对于种间关联显著性的研究,仅用λ2 检验的方法是不够的,因为λ2检验仅得出了关联性显著与否的结论,对那些经检验不显著的种对间,并不意味着它们之间不存在联结性;此外, λ2检验不能区分联结强度的大小,模糊了种间联结性之间的差异,而以λ2检验为基础,结合联结系数AC和共同出现百分率PC共同来测定种对间的联结状况则效果较好。
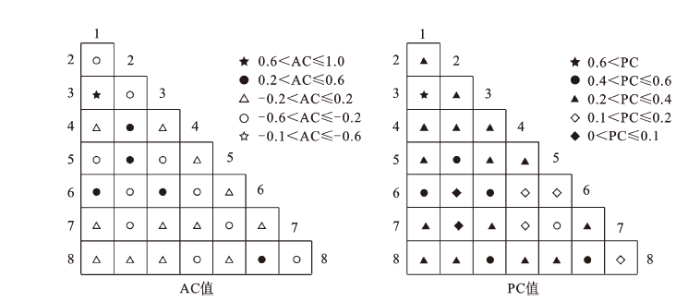
长白山苔原带优势种联结度不强,仅有1对牛皮杜鹃-小白花地榆为强正联结,共同出现的百分率达到60%以上。5对弱正联结, 12对为不联结,10对为弱负联结;5对的共同出现率在40%~60%之间,其余15个种对的共同出现率在20%~40%之间,6个种对共同出现率低于20%(图1)。
图1 优势种群种间联结性半矩阵图
注:1. 牛皮杜鹃 2. 大白花地榆 3. 小白花地榆 4. 小叶章 5. 单花橐吾 6. 高岭风毛菊7. 笃斯越橘 8. 长白老鹳草。
Fig.1 Semi-matrix figure of interspecific association for dominant species
1) 目前长白山苔原带8个优势种中,草本植物已占6个。按重要值分别为牛皮杜鹃、大白花地榆、小白花地榆、小叶章、单花橐吾、高岭风毛菊、笃斯越橘、长白老鹳草。灌木中虽然牛皮杜鹃依然为最主要的优势种,但是笃斯越橘的优势地位已经下降,小白花地榆、大白花地榆和小叶章等多种草本植物已经占据重要位置,长白山苔原植被存在草甸化趋势。
2) 牛皮杜鹃多度大,聚集程度弱,仍为苔原带的植物本底,占据多数生境;笃斯越橘多度小、聚集强度最强,生存空间高度减少,但在部分生境中生长良好;小白花地榆多度较大,聚集程度较弱,与牛皮杜鹃为强正联结,成为强烈扩展的草本植物。大白花地榆、小叶章等草本植物聚集程度较强,呈现斑块分布,已占据了部分生境。多数优势植物之间关联度低,聚集程度较强,说明各优势植物存在生境选择差异,种间竞争导致分布变化的压力较少,苔原带植被变化趋于稳定。
3) 各草本植物入侵、定植、扩展机制多样,依据多度和K值分析,小白花地榆分布广,聚集程度较弱,应为种子繁殖为主,扩展速度应较快;大白花地榆、小叶章等以无性繁殖为主,扩展速度应较慢,但种间竞争能力强,在适应生境中,能有效驱除其它物种。
4) 除牛皮杜鹃与大白花地榆之间存在较显著的负联结,排他现象明显外,多种优势种之间存在正联结,有弱的伴生现象,牛皮杜鹃和小白花地榆为强正联结,伴生现象更明显,可见草本植物与灌木伴生将长期存在。
1) 长白山的高山苔原向高山草甸转化,可和已有的研究比较[18~21]。如在模拟增温和增加氮沉降的条件下,挪威南部高山带的优势矮灌木宽叶仙女木被禾本科草本取代。Bobbink对瑞典北方的亚高山苔原进行氮沉降实验,苔原优势种小灌木岩高兰(Empetrum hermaphroditum)被快速生长的草本曲芒发草(Deschampsia flexuosa)代替。可见,增温和氮沉降增加可能是长白山的高山苔原向高山草甸转化的重要原因。
2) 已有的氮沉降研究表明,氮沉降量增加对入侵的草本植物生长有明显的促进作用,抑制了苔原原生植物特别是笃斯越橘等灌木的生长;在不同氮沉降量条件下,无草本侵入的牛皮杜鹃斑块和笃斯越橘斑块群落结构变化并不明显[11,22]。本研究的结果也表明牛皮杜鹃仍为苔原带的植物本底,笃斯越橘生存空间高度减少。因此,长白山的高山苔原向高山草甸转化过程,可能是缓慢而复杂的。牛皮杜鹃具有较强的抵抗草本植物竞争的能力,而笃斯越橘的竞争能力差[22],可能最先退出苔原带。
3)草本植物聚集程度强,呈斑块状分布。多种草本植物上侵到苔原带,分属不同的植物功能群类型,扩展机制不同,占据不同的空间位置。
草本植物空间格局类型的多样化,是响应苔原带环境变化多样化的结果[23~29],今后的研究要将多样的苔原带植被变化置于统一的全球变化响应模式中,探讨长白山苔原带草本植物空间扩展机理,揭示全球变化下高山苔原带植被变化机理、空间变化过程和未来变化趋势。
The authors have declared that no competing interests exist.
| [1] |
长白山地区近50年平均最高和最低气温变化 [J].https://doi.org/10.3321/j.issn:1000-1522.2009.02.008 URL [本文引用: 1] 摘要
利用线性趋势法对长白山地区13个气象台站建站以来的逐日温度资料进行分析,确定该区域的气温变化趋势,同时通过Mann-Kendall方法确定突变点位置,并结合t检验法进行检验。结果表明:区域的年平均最高气温和最低气温都有极为显著的增加趋势,分别为每10年增加了0.23和0.46℃(α=0.001);近50年的最高气温和最低气温的时间系列都有一个突变点,其中最高气温的突变点出现在1995年左右,最低气温的突变点在1985年前后。这种变化使得温度日较差表现出明显的减小趋势,并在1972年出现一个突变点,前后平均相差0.8℃。区域各月最高、最低气温及气温日较差与日照百分率均有极为显著的相关关系,一定程度上反映了温室效应持续增强的迹象。
Changes in the mean maximum and minimum temperatures in Changbai Mountain,northeastern China in the past 50 years. https://doi.org/10.3321/j.issn:1000-1522.2009.02.008 URL [本文引用: 1] 摘要
利用线性趋势法对长白山地区13个气象台站建站以来的逐日温度资料进行分析,确定该区域的气温变化趋势,同时通过Mann-Kendall方法确定突变点位置,并结合t检验法进行检验。结果表明:区域的年平均最高气温和最低气温都有极为显著的增加趋势,分别为每10年增加了0.23和0.46℃(α=0.001);近50年的最高气温和最低气温的时间系列都有一个突变点,其中最高气温的突变点出现在1995年左右,最低气温的突变点在1985年前后。这种变化使得温度日较差表现出明显的减小趋势,并在1972年出现一个突变点,前后平均相差0.8℃。区域各月最高、最低气温及气温日较差与日照百分率均有极为显著的相关关系,一定程度上反映了温室效应持续增强的迹象。
|
| [2] |
1959-2006年长白山地区降水序列的多时间尺度分析 [J].Magsci 摘要
<p>基于长白山地区松江、东岗、长白、和龙、临江和天池6个气象站1959—2006年的月均降水量和年降水量数据,采用Morlet小波分析方法,对1959—2006年长白山地区植被生长季(5—9月)降水量、降雪季(11月至次年4月)降水量和年降水量序列进行多尺度特征分析,并运用Daubechies小波系中的db5小波对各降水序列进行不同层次的分解和低频重构,对重构序列进行了趋势识别和分析.结果表明:研究期间,长白山地区植被生长季降水量存在3~6 a、10~13 a和24~30 a的特征周期;降雪季降水量存在1~2 a、5~7 a和17~20 a的特征周期;年降水存在8~10 a、16~20 a、25~30 a的特征周期;研究区年降水量序列呈现整体下降的趋势.</p>
Multiple time scale analysis of precipitation series in Changbai Mountain Region in 1959-2006. Magsci 摘要
<p>基于长白山地区松江、东岗、长白、和龙、临江和天池6个气象站1959—2006年的月均降水量和年降水量数据,采用Morlet小波分析方法,对1959—2006年长白山地区植被生长季(5—9月)降水量、降雪季(11月至次年4月)降水量和年降水量序列进行多尺度特征分析,并运用Daubechies小波系中的db5小波对各降水序列进行不同层次的分解和低频重构,对重构序列进行了趋势识别和分析.结果表明:研究期间,长白山地区植被生长季降水量存在3~6 a、10~13 a和24~30 a的特征周期;降雪季降水量存在1~2 a、5~7 a和17~20 a的特征周期;年降水存在8~10 a、16~20 a、25~30 a的特征周期;研究区年降水量序列呈现整体下降的趋势.</p>
|
| [3] |
Climate variability,ecological gradient and the Northeast China Transect(NECT) [J].https://doi.org/10.1006/jare.2000.0667 URL 摘要
The general characteristics of the NECT are presented and the interannual and innerannual variability of temperature, precipitation, potential evapotranspiration and potential evapotranspiration ratio were studied to describe current levels of temporal and spatial variability in the NECT climate. Important factors in determining the structure and function of NECT ecosystems as well as the tempo...
|
| [4] |
近52a长白山苔原带生长季气候变化特征 [J].Magsci 摘要
利用长白山苔原带天池站1959—2010年6—9月气象数据,进行生长季气候变化分析。采用距平值、小波分析、MannKendall检验相结合的方法,进行温度、降水周期性变化与突变点的检验;采用线性倾向估计法,分析未来气候变化的趋势;并通过气候变化指数,反映其气候变化情况。结果显示:52 a来,长白山苔原带生长季经历了暖湿—冷干—暖湿—冷干—冷湿—冷干—暖干几个主要阶段,温度变化周期为5~10 a,降水变化周期为5 a和20 a.。温度变化的突变年份发生在1998年,降水变化较之温度变化复杂,其存在多个突变年份;各气象要素的变化趋势大都表现为显著的升高,其中温度的变化最为明显,风速、日照时数的变化甚微;霜冻日数与冰点日数均显著减少,极端温度大多出现在21世纪;伴随着>0.1 mm降雨日数的显著减小,长白山苔原带降水强度不断增加;与植物生长相关的生长季≥0 ℃积温、≥3 ℃积温和≥5 ℃积温在52 a间呈显著增加趋势,积温的增加将会对生长季的延长及植物生长起到显著的促进作用。
Study on climate change in alpine tundra of the Changbai Mountain in growing season in recent 52 years. Magsci 摘要
利用长白山苔原带天池站1959—2010年6—9月气象数据,进行生长季气候变化分析。采用距平值、小波分析、MannKendall检验相结合的方法,进行温度、降水周期性变化与突变点的检验;采用线性倾向估计法,分析未来气候变化的趋势;并通过气候变化指数,反映其气候变化情况。结果显示:52 a来,长白山苔原带生长季经历了暖湿—冷干—暖湿—冷干—冷湿—冷干—暖干几个主要阶段,温度变化周期为5~10 a,降水变化周期为5 a和20 a.。温度变化的突变年份发生在1998年,降水变化较之温度变化复杂,其存在多个突变年份;各气象要素的变化趋势大都表现为显著的升高,其中温度的变化最为明显,风速、日照时数的变化甚微;霜冻日数与冰点日数均显著减少,极端温度大多出现在21世纪;伴随着>0.1 mm降雨日数的显著减小,长白山苔原带降水强度不断增加;与植物生长相关的生长季≥0 ℃积温、≥3 ℃积温和≥5 ℃积温在52 a间呈显著增加趋势,积温的增加将会对生长季的延长及植物生长起到显著的促进作用。
|
| [5] |
2000-2008年长白山地区植被覆盖变化对气候的响应研究 [J].URL 摘要
利用2000~2008年MODIS/NDVI数据,结合谐波分析、影像处理和基于像元的空间统计方法,分析了不同时间尺度下长白山地区的植被覆盖年内和年际变化与气温、降水的空间相关性。结果表明,近10a来长白山地区气温和降水都均呈增加趋势;年内和年际变化过程中,长白山地区植被覆盖受气温影响的程度要高于降水;长白山地区春季和秋季植被覆盖与气温呈正相关,夏季主要呈负相关;不同月份的相关统计与年统计和分季节统计相比,更能细致地反映植被覆盖与气候的响应关系。
The responses of vegetation cover to climate change in the Changbai Mountain area from 2000 to 2008. URL 摘要
利用2000~2008年MODIS/NDVI数据,结合谐波分析、影像处理和基于像元的空间统计方法,分析了不同时间尺度下长白山地区的植被覆盖年内和年际变化与气温、降水的空间相关性。结果表明,近10a来长白山地区气温和降水都均呈增加趋势;年内和年际变化过程中,长白山地区植被覆盖受气温影响的程度要高于降水;长白山地区春季和秋季植被覆盖与气温呈正相关,夏季主要呈负相关;不同月份的相关统计与年统计和分季节统计相比,更能细致地反映植被覆盖与气候的响应关系。
|
| [6] |
长白山岳桦苔原过渡带动态与气候变化 [J].https://doi.org/10.3321/j.issn:1005-2321.2002.01.027 URL 摘要
应用样带网格调查方法 ,对长白山岳桦苔原过渡带上岳桦径级结构、分布格局以及过渡带的环境因子进行调查 ,并与最近几十年的极端低温进行了对比研究 ,用格局的差异和径级结构的动态变化来揭示全球变暖对岳桦苔原过渡带的影响。结果表明 :全球气候变暖对长白山岳桦苔原过渡带产生很大影响 ,气候变暖使得岳桦苔原过渡带中的岳桦分布主要以幼苗和幼树为主 ,其中岳桦的径级分布呈倒J字型 ,过渡带中岳桦种群格局呈聚集型分布。整个岳桦种群随着全球气候变暖有一种整体向上迁移的趋势 ,尤其以岳桦苔原过渡带最为明显 ,岳桦苔原过渡带变宽 ,岳桦向苔原侵入的程度加剧。
The effect of global climate change on the dynamics of Betula ermanii-tundra ecotone in the Changbai Mountains. https://doi.org/10.3321/j.issn:1005-2321.2002.01.027 URL 摘要
应用样带网格调查方法 ,对长白山岳桦苔原过渡带上岳桦径级结构、分布格局以及过渡带的环境因子进行调查 ,并与最近几十年的极端低温进行了对比研究 ,用格局的差异和径级结构的动态变化来揭示全球变暖对岳桦苔原过渡带的影响。结果表明 :全球气候变暖对长白山岳桦苔原过渡带产生很大影响 ,气候变暖使得岳桦苔原过渡带中的岳桦分布主要以幼苗和幼树为主 ,其中岳桦的径级分布呈倒J字型 ,过渡带中岳桦种群格局呈聚集型分布。整个岳桦种群随着全球气候变暖有一种整体向上迁移的趋势 ,尤其以岳桦苔原过渡带最为明显 ,岳桦苔原过渡带变宽 ,岳桦向苔原侵入的程度加剧。
|
| [7] |
长白山北坡林线岳桦种群动态对气候变化响应的坡向分异 [J].URL 摘要
选取长白山北坡岳桦(Betula ermanii)苔原交错带3个不同坡向(东北坡、东南坡、东坡)的岳桦径级结构为研究对象,分析坡向在岳桦种群动态与气候变化之间关系的异同。结果表明:三坡向的岳桦种群扩张和气温高低存在明显差异。14个生态气候指标与对应年份内繁殖且存活下来的岳桦棵数的逐步回归分析和相关分析表明三坡向岳桦种群扩张是对气温变化的积极响应,但响应有差异:东坡东南坡东北坡。不同坡向岳桦种群扩张的差异是对气温变化差异的响应。
The dynamics response of betula ermanii population and climate change on different slopes aspect of North slope, changbai mountains. URL 摘要
选取长白山北坡岳桦(Betula ermanii)苔原交错带3个不同坡向(东北坡、东南坡、东坡)的岳桦径级结构为研究对象,分析坡向在岳桦种群动态与气候变化之间关系的异同。结果表明:三坡向的岳桦种群扩张和气温高低存在明显差异。14个生态气候指标与对应年份内繁殖且存活下来的岳桦棵数的逐步回归分析和相关分析表明三坡向岳桦种群扩张是对气温变化的积极响应,但响应有差异:东坡东南坡东北坡。不同坡向岳桦种群扩张的差异是对气温变化差异的响应。
|
| [8] |
长白山北坡岳桦林线变动的水热条件分析 [J].https://doi.org/10.11820/dlkxjz.2011.03.008 Magsci 摘要
近50 年来长白山北坡林线种群呈现明显的扩张态势。为揭示林线变动机制,本文以长白山天池气象站的气象数据(1953-2007 年)为基础,结合野外的气温观测,以温暖指数(<em>WI</em>)及湿润指数(<em>HI</em>)为生态气候指标,以16℃·月和68.8 mm/℃·月为阈值,通过数据保证率的计算推测55 年来长白山北坡岳桦林线的水热条件变化。结果表明:以<em>WI</em> 和<em>HI</em> 指标确定的林线位置分别在1975 ~ 2460 m和1584 ~ 2231 m。WI 与HI 变化对林线高度变动影响的交互作用不显著(p>0.05),<em>WI</em> 与<em>HI</em> 变化呈显著负相关(<em>p</em><0.01),两种林线变化也呈负相关(<em>r</em>=-0.11<0),交互作用和相关分析结果显示水热条件的不同步变化使岳桦林线上侵不能达到各自的潜在高度,林线位置波动在1975 ~ 2231 m。林线上缘波动的复杂程度高于下缘,波动幅度与胁迫力的大小呈正相关。
Water and heat changes of Betula ermanii treeline on northern slope of Changbai Mountains. https://doi.org/10.11820/dlkxjz.2011.03.008 Magsci 摘要
近50 年来长白山北坡林线种群呈现明显的扩张态势。为揭示林线变动机制,本文以长白山天池气象站的气象数据(1953-2007 年)为基础,结合野外的气温观测,以温暖指数(<em>WI</em>)及湿润指数(<em>HI</em>)为生态气候指标,以16℃·月和68.8 mm/℃·月为阈值,通过数据保证率的计算推测55 年来长白山北坡岳桦林线的水热条件变化。结果表明:以<em>WI</em> 和<em>HI</em> 指标确定的林线位置分别在1975 ~ 2460 m和1584 ~ 2231 m。WI 与HI 变化对林线高度变动影响的交互作用不显著(p>0.05),<em>WI</em> 与<em>HI</em> 变化呈显著负相关(<em>p</em><0.01),两种林线变化也呈负相关(<em>r</em>=-0.11<0),交互作用和相关分析结果显示水热条件的不同步变化使岳桦林线上侵不能达到各自的潜在高度,林线位置波动在1975 ~ 2231 m。林线上缘波动的复杂程度高于下缘,波动幅度与胁迫力的大小呈正相关。
|
| [9] |
Responses of tree islands to air temperature change at treeline on north-facing slopes of the Changbai Mountains [J].https://doi.org/10.2747/0272-3646.32.4.374 Magsci 摘要
A total of 60 scattered tree islands from seven typical plots were collected in the upper treeline on the north-facing slopes of the Changbai Mountains. We selected eight ecological variables related to air temperature, which were simulated based on measured air temperature data and the records at the Tianchi meteorological station. Annual stem recruitment of the Betula ermanii (Ermans Birch) population was calculated using tree-ring analysis and field observations. We linked these ecological variables with annual stem recruitment from 1953 to 2008 to examine the relationship between population dynamics and air temperature change. Our results showed that the mean annual rate of air temperature increase was 0.045 degrees C/yr between 1953 and 2008, which was far higher than that of global average warming, suggesting that the study area may be more sensitive and vulnerable to climate change. Different propagation forms and annual stem recruitment of the Betula ermanii population responded significantly to the increase of air temperature. The population first occupied a point niche using the shrub life form during the period 1953-1988, when air temperature increased at a slow pace (0.007 degrees C/yr on average). In contrast, the population expanded its niche rapidly using a single-trunk life form during the period 1988-2008, when air temperature increased at a much faster pace (0.11 degrees C/yr on average). Under global warming, the population expansion and the domain enlargement of Betula ermanii led to a treeline shift. The population has continued to vary its physiological traits to adapt to air temperature change.
|
| [10] |
Current and potential tree locations in tree line ecotone of Changbai Mountains, Northeast China: the controlling effects of topography [J]. |
| [11] |
Effects of Nitrogen deposition on tundra vegetation undergoing invasion by Deyeuxia angustifolia in Changbai Mountains [J].https://doi.org/10.1007/s11769-015-0746-1 URL [本文引用: 2] 摘要
在最近的年里,草本的种类象 Deyeuxia angustifolia (Kom ) 那样 Y. L。Chang 入侵了 Changbai 山的西方的斜坡的高山的冻土带区域。因为大气的氮免职被预言在温暖的气候和 D 下面增加。angustifolia 对氮增加敏感,地实验从 2010 ~ 2013 被进行在 D 的机制上决定增加的氮免职的效果。angustifolia 侵略。这研究的目标是在高山的冻土带植被在变化上评估增加的氮免职的影响(主要由杜鹃花属 chrysanthum 组成棺罩。并且 Vaccinium uliginosum 悬崖) 。结果显示出那:1 ) 模仿的氮免职影响了 R 的全面特征和结构。chrysanthum 和 V。uliginosum 社区并且冻土带植被的生长上的积极影响由 D 入侵了。angustifolia;2 ) R。chrysanthum 对由 D 的侵略更抵抗。angustifolia 比 V。uliginosum;3 ) 模仿的氮免职能改进生长并且提高 D 的竞争力。angustifolia,它是逐渐地代替的 R。chrysanthum 和 V。uliginosum 和力量在未来在系统成为主导的种类,把高山的冻土带转变成在 Changbai 山的高山的草地。
|
| [12] |
长白山高山冻原植物的调查研究简报 [J].
长白山是东北亚大陆上唯一的、最高的具有原始高山冻原植被的名山。位于北纬41°23′——42°36′,东径126°55′——129°。这一大片经过多次多处喷火的火山群,在喷火次数最多最集中的地方形成了长白山的主体,即白头山。因其山势雄伟、资源丰富,早为国内外有关学科的工作者们所向往。在中国侧自1886年起单就从事植物学调查采集的主要工作者们已有10几位。但专门对高山冻原植物的研究报告却很少。只见到竹内亮教授的手稿。目前多数同志们认为在海拔2000——2100公尺以上是高山冻原带;在海拔2300——2400公尺处可分为两个亚带,并把下部的叫做“高山常绿矮石楠灌丛”;上部的叫做“高山半荒漠”。他们又作了 Raunkiar 氏生活型谱及小群丛嵌复合体的文字和图表的分析描述。“附录”是高山带上现生已知的植物名录。
A brief report on the research of the Changbaishan alpine tundra vegetation.
长白山是东北亚大陆上唯一的、最高的具有原始高山冻原植被的名山。位于北纬41°23′——42°36′,东径126°55′——129°。这一大片经过多次多处喷火的火山群,在喷火次数最多最集中的地方形成了长白山的主体,即白头山。因其山势雄伟、资源丰富,早为国内外有关学科的工作者们所向往。在中国侧自1886年起单就从事植物学调查采集的主要工作者们已有10几位。但专门对高山冻原植物的研究报告却很少。只见到竹内亮教授的手稿。目前多数同志们认为在海拔2000——2100公尺以上是高山冻原带;在海拔2300——2400公尺处可分为两个亚带,并把下部的叫做“高山常绿矮石楠灌丛”;上部的叫做“高山半荒漠”。他们又作了 Raunkiar 氏生活型谱及小群丛嵌复合体的文字和图表的分析描述。“附录”是高山带上现生已知的植物名录。
|
| [13] |
长白山高山冻原植物群落的数量分类和排序 [J].
本文用目前在国际上比较先进的双向指示种分析(TWINSPAN)多元等级分划分类方法和无趋势对应分析(DCA)排序方法对长自山高山冻原植物群落进行了研究。研究结果表明:DCA的第2轴与长白山高山冻原59个植物群落类型的土壤水分梯度紧密相关, TWINSPAN将长白山高山冻原59个植物群落分为12组。此外, 本文还对长白山高山冻原植物群落的TWINSPAN和DCA分类结果与经典分类方法对其分类的结果进行了比较。
Numerical classification and ordination of plant communities in Mt.Changbai.
本文用目前在国际上比较先进的双向指示种分析(TWINSPAN)多元等级分划分类方法和无趋势对应分析(DCA)排序方法对长自山高山冻原植物群落进行了研究。研究结果表明:DCA的第2轴与长白山高山冻原59个植物群落类型的土壤水分梯度紧密相关, TWINSPAN将长白山高山冻原59个植物群落分为12组。此外, 本文还对长白山高山冻原植物群落的TWINSPAN和DCA分类结果与经典分类方法对其分类的结果进行了比较。
|
| [14] |
|
| [15] |
火山干扰下的长白山植被分布规律 [J].Magsci 摘要
<p>距今约800 a前,长白山天池火山发生大规模普林尼式喷发,喷出的巨量火山浮石及火山灰毁灭性地破坏了长白山植被。由于受喷发倾角和强劲高空西北风影响,使各坡向火山灰和浮石的沉积厚度不同,东坡最厚,南坡和西坡次之,北坡最少。火山灰与浮石形成疏松的成土母质,在流水持续侵蚀下,对破坏后的植被演替产生了长期影响。利用野外调查和现有资料,论证了长白山植被分布独特性规律是长白山火山喷发及喷发物水蚀过程对生态系统作用的结果,确认火山喷发虽历经800多a,目前各坡向植被仍然处在演替的不同进程之中,呈现差异性分布规律。</p>
Effects of volcanic interference on the vegetation distribution of Changbai Mountain. Magsci 摘要
<p>距今约800 a前,长白山天池火山发生大规模普林尼式喷发,喷出的巨量火山浮石及火山灰毁灭性地破坏了长白山植被。由于受喷发倾角和强劲高空西北风影响,使各坡向火山灰和浮石的沉积厚度不同,东坡最厚,南坡和西坡次之,北坡最少。火山灰与浮石形成疏松的成土母质,在流水持续侵蚀下,对破坏后的植被演替产生了长期影响。利用野外调查和现有资料,论证了长白山植被分布独特性规律是长白山火山喷发及喷发物水蚀过程对生态系统作用的结果,确认火山喷发虽历经800多a,目前各坡向植被仍然处在演替的不同进程之中,呈现差异性分布规律。</p>
|
| [16] |
长白山西坡小叶章侵入苔原带调查与机理分析 [J].https://doi.org/10.3969/j.issn.1008-2786.2013.04.010 URL [本文引用: 1] 摘要
受火山活动干扰,长白山各坡向植被演替进程表现出较大差异,东坡植被演替处于早期阶段;而北坡出现岳桦(BetulaermaniiCharm.)上侵现象;南坡植被演替处于二者之间。经过多年野外调查对比分析,首次提出长白山西坡苔原带出现小叶章(Deyeuxia.angustifolia(Kom.)Y.L.Chang)侵入苔原的现象。调查显示小叶章呈现从低海拔处向高海拔逐渐侵入的趋势,威胁牛皮杜鹃(RhododendronchrysanthumPall.)及笃斯越橘(Vacciniumuligino—sumLinn.)等苔原带植物的生存,并导致苔原带景观变化。在调查长白山西坡小叶章侵入苔原带的基础上,根据苔原带的环境变化设计了小叶章移栽与增温实验、小叶章种子着床实验。实证了小叶章上侵是火山喷发后植被演替的一个特殊阶段,并提出长白山苔原带西坡小叶章上侵代表了岳桦林带上移的理论。
Investigation and mechanism analysis on the invasion of deyeuxia.angustifolia to tundra Zone in western slope of changbai mountain. https://doi.org/10.3969/j.issn.1008-2786.2013.04.010 URL [本文引用: 1] 摘要
受火山活动干扰,长白山各坡向植被演替进程表现出较大差异,东坡植被演替处于早期阶段;而北坡出现岳桦(BetulaermaniiCharm.)上侵现象;南坡植被演替处于二者之间。经过多年野外调查对比分析,首次提出长白山西坡苔原带出现小叶章(Deyeuxia.angustifolia(Kom.)Y.L.Chang)侵入苔原的现象。调查显示小叶章呈现从低海拔处向高海拔逐渐侵入的趋势,威胁牛皮杜鹃(RhododendronchrysanthumPall.)及笃斯越橘(Vacciniumuligino—sumLinn.)等苔原带植物的生存,并导致苔原带景观变化。在调查长白山西坡小叶章侵入苔原带的基础上,根据苔原带的环境变化设计了小叶章移栽与增温实验、小叶章种子着床实验。实证了小叶章上侵是火山喷发后植被演替的一个特殊阶段,并提出长白山苔原带西坡小叶章上侵代表了岳桦林带上移的理论。
|
| [17] |
长白山西坡小叶章侵入苔原带过程及影响研究 [J].https://doi.org/10.5846/stxb201302270319 URL Magsci [本文引用: 1] 摘要
长白山西坡岳桦林带的草本植物(以小叶章为代表)侵入了苔原带,形成了独特的植物入侵现象。在光谱及影像分析的基础上,结合GPS(Global Positional System)定位技术,并依据小叶章与牛皮杜鹃的光谱差异及其反演的NDVI (Normalized Difference Vegetation Index)植被指数,揭示小叶章侵入苔原带的过程;通过对不同侵入时间、强度的斑块进行群落调查及土壤测试,探究小叶章侵入苔原带的生态后果。结果显示小叶章侵入苔原带始于20世纪80年代后期,由低海拔向高海拔推进,进入21世纪后逐渐形成了稳定的以小叶章为优势物种的植物群落结构。目前,低海拔处的小叶章斑块经过多年扩张已连接成片,而高海拔处的斑块正处于扩张的初期阶段。从生物多样性变化可以看出,小叶章侵入苔原带导致植物群落多样性升高和物种数量的增加,苔原带原有的灌木数量明显减少,草本植物逐渐增多。植被的改变影响了土壤的理化性质,C/N比下降,土壤腐殖质含量和全氮含量下降,但速效氮和土壤持水能力上升,土壤养分的高效利用又进一步推动了小叶章的侵入。小叶章侵入苔原带已经造成了严重的生态后果。
Analysis on the process and impacts of deyeuxia angustifolia invasion on the alpine tundra, Changbai Mountain. https://doi.org/10.5846/stxb201302270319 URL Magsci [本文引用: 1] 摘要
长白山西坡岳桦林带的草本植物(以小叶章为代表)侵入了苔原带,形成了独特的植物入侵现象。在光谱及影像分析的基础上,结合GPS(Global Positional System)定位技术,并依据小叶章与牛皮杜鹃的光谱差异及其反演的NDVI (Normalized Difference Vegetation Index)植被指数,揭示小叶章侵入苔原带的过程;通过对不同侵入时间、强度的斑块进行群落调查及土壤测试,探究小叶章侵入苔原带的生态后果。结果显示小叶章侵入苔原带始于20世纪80年代后期,由低海拔向高海拔推进,进入21世纪后逐渐形成了稳定的以小叶章为优势物种的植物群落结构。目前,低海拔处的小叶章斑块经过多年扩张已连接成片,而高海拔处的斑块正处于扩张的初期阶段。从生物多样性变化可以看出,小叶章侵入苔原带导致植物群落多样性升高和物种数量的增加,苔原带原有的灌木数量明显减少,草本植物逐渐增多。植被的改变影响了土壤的理化性质,C/N比下降,土壤腐殖质含量和全氮含量下降,但速效氮和土壤持水能力上升,土壤养分的高效利用又进一步推动了小叶章的侵入。小叶章侵入苔原带已经造成了严重的生态后果。
|
| [18] |
Simulated climate change altered dominance hierarchies and diversity of an alpine biodiversity hotspot [J].https://doi.org/10.1890/04-1563 URL [本文引用: 1] 摘要
Alpine and arctic ecosystems may be particularly vulnerable to climate change. We know little about alpine plant community responses to the predicted abiotic changes, or to possible changes in the biotic environment caused by climate change. Four years of experimental warming and nutrient addition altered dominance hierarchies, community structure, and diversity of an alpine biodiversity hotspot in south Norway. The previously dominant dwarf shrub Dryas octopetala was replaced by graminoids and forbs under nutrient addition and warming with nutrients. Community diversity declined due to decreased bryophyte and lichen richness and abundances, and dwarf shrub abundances. In controls and in plots with only warming, where Dryas maintained dominance, the relationships between changes in Dryas cover and changes in community parameters were negative, suggesting that Dryas controls community processes. Under nutrient addition, bryophyte and lichen diversity decreased with decreasing Dryas cover, probably due to i...
|
| [19] |
The effects of air-borne Nitrogen pollutants on species diversity in natural and semi-natural European vegetation [J].https://doi.org/10.1046/j.1365-2745.1998.8650717.x URL 摘要
Summary The effects of increased atmospheric nitrogen inputs, from both NO y and NH x, on diversity in various semi-natural and natural ecosystems are reviewed. The severity of these impacts depends on abiotic conditions (e.g. buffering capacity, soil nutrient status and soil factors that influence the nitrification potential and nitrogen immobilization rate) in the particular system. The sensitivity of fresh water ecosystems, wetlands and bogs, species-rich grasslands, heathlands and field layer of forests, all of which have conservational value, are discussed in detail. The most important effects of nitrogen deposition are: (i) accumulation of nitrogenous compounds resulting in enhanced availability of nitrate or ammonium; (ii) soil-mediated effects of acidification; and (iii) increased susceptibility to secondary stress factors. Long-term nitrogen enrichment has gradually increased the availability of nitrogen in several vegetation types, leading to competitive exclusion of characteristic species by more nitrophilic plants, especially under oligo- to mesotrophic soil conditions. Soil acidification (with losses of buffering capacity and increased concentrations of toxic metals) is especially important after nitrification of ammonium in weakly buffered environments: acid-resistant plant species then become dominant at the expense of the often rare plants typical of intermediate pH. The related change in the balance between ammonium and nitrate may also affect the performance of several species. The susceptibility of plant species to secondary stress factors (pathogens; frost and drought) may be affected by air-borne nitrogen but data are only available for a few communities (e.g. dry heathlands). Most global biodiversity is contained within natural and semi-natural vegetation. It is thus crucial to control emissions of nitrogenous compounds to the atmosphere, in order to reduce or prevent effects on diversity in these systems. Most research has focused on acidification in forestry stands and lakes and on the effects on trees. We highlight serious gaps in knowledge of other ecosystems.
|
| [20] |
The relative role of dispersal and local interactions for alpine plant community diversity under simulated climate warming [J].https://doi.org/10.1111/j.0030-1299.2007.15906.x URL 摘要
Most studies on factors determining diversity are conducted in temperate or warm regions, whereas studies in climatically harsh and low productivity areas, such as alpine regions, are rare. We examined the relative roles of seed availability and different biotic and abiotic factors for the diversity of an alpine plant community in southern Norway. Furthermore, because climate warming is predicted to be an important driver of alpine species diversity, we assessed how the relative impacts of dispersal and local interactions on diversity might change under experimental warming (open top chambers, OTCs).Addition of seeds from 27 regional species increased community diversity. The establishment of the species was negatively related both to the diversity of the existing system and the cover of the abundant dwarf shrub . These results show that both species dispersal limitation and local biotic interactions are important factors for alpine plant community diversity. Despite relatively harsh environmental conditions and low productivity, competition from the resident vegetation appeared to have a greater role for species establishment and diversity than facilitation and experimental warming. Higher temperature appeared to increase the negative relationship between resident species diversity and species establishment. This may suggest that climate warming can increase the role of interspecific competition for alpine plant community structure, and thus alter the long-term effects of biotic interactions on diversity.
|
| [21] |
Species-specific responses of an alpine plant community under simulated environmental change [J]. |
| [22] |
氮沉降对长白山苔原植被影响的试验研究 [J].
<p>为了探讨氮沉降增加对苔原植被的影响,特别是对草本植物侵入苔原的作用,在长白山高山苔原带进行了连续4a的人工氮沉降模拟实验,测定3种设定的氮沉降水平下牛皮杜鹃(<em>Rhododendron chrysanthum</em>)、笃斯越橘(<em>Vaccinium uliginosum</em>)和小叶章(<em>Deyeuxia angustifolia.</em>)的生长状况和群落结构变化.研究结果表明:① 与牛皮杜鹃、笃斯越橘等苔原本地种相比,氮沉降量增加更有利于侵入的小叶章生长;牛皮杜鹃生长对氮沉降量变化响应微弱,氮沉降量增加能明显抑制笃斯越橘的生长.② 无小叶章侵入的牛皮杜鹃和笃斯越橘斑块在不同氮沉降量条件下,群落结构变化不明显,氮沉降增加不是小叶章侵入苔原带的直接原因.③ 在小叶章侵入牛皮杜鹃和笃斯越橘斑块后,氮沉降量增加强化了小叶章的竞争能力,逐渐取代牛皮杜鹃或笃斯越橘,成为优势种,推动高山苔原向高山草甸转化.因此,随着氮沉降量的不断增加,长白山苔原带将面临退化与草甸化.</p>
Zong Shengwei
<p>为了探讨氮沉降增加对苔原植被的影响,特别是对草本植物侵入苔原的作用,在长白山高山苔原带进行了连续4a的人工氮沉降模拟实验,测定3种设定的氮沉降水平下牛皮杜鹃(<em>Rhododendron chrysanthum</em>)、笃斯越橘(<em>Vaccinium uliginosum</em>)和小叶章(<em>Deyeuxia angustifolia.</em>)的生长状况和群落结构变化.研究结果表明:① 与牛皮杜鹃、笃斯越橘等苔原本地种相比,氮沉降量增加更有利于侵入的小叶章生长;牛皮杜鹃生长对氮沉降量变化响应微弱,氮沉降量增加能明显抑制笃斯越橘的生长.② 无小叶章侵入的牛皮杜鹃和笃斯越橘斑块在不同氮沉降量条件下,群落结构变化不明显,氮沉降增加不是小叶章侵入苔原带的直接原因.③ 在小叶章侵入牛皮杜鹃和笃斯越橘斑块后,氮沉降量增加强化了小叶章的竞争能力,逐渐取代牛皮杜鹃或笃斯越橘,成为优势种,推动高山苔原向高山草甸转化.因此,随着氮沉降量的不断增加,长白山苔原带将面临退化与草甸化.</p>
|
| [23] |
Biotic interactions limit species richness in an alpine plant community, especially under experimental warming [J].https://doi.org/10.1111/j.1600-0706.2013.00336.x Magsci [本文引用: 1] 摘要
The determinants of local species richness in plant communities have been the subject of much debate. Is species richness the result of stochastic events such as dispersal processes, or do local environmental filters sort species into communities according to their ecological niches? Recent studies suggest that these two processes simultaneously limit species richness, although their relative importance may vary in space and time. Understanding the limiting factors for species richness is especially important in light of the ongoing global warming, as new species establish in resident plant communities as a result of climate-driven migration. We examined the relative importance of dispersal and environmental filtering during seedling recruitment and plant establishment in an alpine plant community subjected to seed addition and long-term experimental warming. Seed addition increased species richness during the seedling recruitment stage, but this initial increase was cancelled out by a corresponding decrease in species richness during plant establishment, suggesting that environmental filters limit local species richness in the long term. While initial recruitment success of the sown species was related to both abiotic and biotic factors, long-term establishment was controlled mainly by biotic factors, indicating an increase in the relative importance of biotic interactions once plants have germinated in a microhabitat with favourable abiotic conditions. The relative importance of biotic interactions also seemed to increase with experimental warming, suggesting that increased competition within the resident vegetation may decrease community invasibility as the climate warms.
|
| [24] |
Indirect effects of global change accumulate to alter plant diversity but not ecosystem function in alpine tundra [J]. |
| [25] |
A review of snow manipulation experiments in Arctic and alpine tundra ecosystems [J].https://doi.org/10.3402/polar.v29i1.6054 URL 摘要
Abstract Snow cover is one of the most important factors controlling microclimate and plant growing conditions for Arctic and alpine ecosystems. Climate change is altering snowfall regimes, which in turn influences snow cover and ultimately tundra plant communities. The interest in winter climate change and the number of experiments exploring the responses of alpine and Arctic ecosystems to changes in snow cover have been growing in recent years, but their outcomes are difficult to summarize because of the large variability in manipulation approaches, extents and measured response variables. In this review, we (1) compile the ecological publications on snow manipulation experiments, (2) classify the studies according to the climate scenarios they simulate and response variables they measure, (3) discuss the methods applied to manipulate snow cover, and (4) analyse and generalize the response in phenology, productivity and community composition by means of a meta-analysis. This meta-analysis shows that flowering phenology responded strongly to changes in the timing of snowmelt. The least responsive group of species were graminoids; however, they did show a decrease in productivity and abundance with experimentally increased snow covers. The species group with the greatest phenological response to snowmelt changes were the dwarf shrubs. Their abundance also increased in most long-term snow fence experiments, whereas species richness generally declined. We conclude that snow manipulation experiments can improve our understanding of recently observed ecosystem changes, and are an important component of climate change research.
|
| [26] |
A significant upward shift in plant species optimum elevation during the 20th century [J].https://doi.org/10.1126/science.1156831 URL PMID: 18583610 摘要
Spatial fingerprints of climate change on biotic communities are usually associated with changes in the distribution of species at their latitudinal or altitudinal extremes. By comparing the altitudinal distribution of 171 forest plant species between 1905 and 1985 and 1986 and 2005 along the entire elevation range (0 to 2600 meters above sea level) in west Europe, we show that climate warming has resulted in a significant upward shift in species optimum elevation averaging 29 meters per decade. The shift is larger for species restricted to mountain habitats and for grassy species, which are characterized by faster population turnover. Our study shows that climate change affects the spatial core of the distributional range of plant species, in addition to their distributional margins, as previously reported.
|
| [27] |
Introduction of snow and geomorphic disturbance variables into predictive models of alpine plant distribution in the western Swiss Alps [J].https://doi.org/10.1657/1938-4246-41.3.347 URL 摘要
Indirect topographic variables have been used successfully as surrogates for disturbance processes in plant species distribution models (SDM) in mountain environments. However, no SDM studies have directly tested the performance of disturbance variables. In this study, we developed two disturbance variables: a geomorphic index (GEO) and an index of snow redistribution by wind (SNOW). These were...
|
| [28] |
高山植物对全球气候变暖的响应研究进展 [J].URL 摘要
高山地区被认为是对全球气候变化最敏感的区域之一,气候变暖导致高山生境内生物与非生物环境因子发生不同程度的改变,从而引起高山植物从不同层面作出不同的变化或响应.综述了高山及亚高山高海拔生境内,植物从宏观生态系统层面到微观个体生理层面对全球气候变化进行响应的研究进展.植物由个体自身的生理及形态上产生不同响应,逐步经过"瀑布式上升效应(Cascade effects)",最后引起整个高山生态系统的转变.受局地差异性及物种差异性影响,生物多样性在不同高山地区呈现出或是增加或是减少的趋势;林线及植被向高纬度、高海拔地区扩张;植物间相互关系由协作转向中性乃至竞争;植物物候、繁殖、生物量生产、光合作用、年轮生长、营养结构等方面均呈现出不同的响应模式.这些从微观到宏观的不同响应模式,最终将引起高山生态系统在结构、功能上的改变,进而在很大范围上威胁到高山植物的生存与发展.最后提出该领域未来的研究重点.参147
Advances in the studies of responses of alpine plants to global warming, URL 摘要
高山地区被认为是对全球气候变化最敏感的区域之一,气候变暖导致高山生境内生物与非生物环境因子发生不同程度的改变,从而引起高山植物从不同层面作出不同的变化或响应.综述了高山及亚高山高海拔生境内,植物从宏观生态系统层面到微观个体生理层面对全球气候变化进行响应的研究进展.植物由个体自身的生理及形态上产生不同响应,逐步经过"瀑布式上升效应(Cascade effects)",最后引起整个高山生态系统的转变.受局地差异性及物种差异性影响,生物多样性在不同高山地区呈现出或是增加或是减少的趋势;林线及植被向高纬度、高海拔地区扩张;植物间相互关系由协作转向中性乃至竞争;植物物候、繁殖、生物量生产、光合作用、年轮生长、营养结构等方面均呈现出不同的响应模式.这些从微观到宏观的不同响应模式,最终将引起高山生态系统在结构、功能上的改变,进而在很大范围上威胁到高山植物的生存与发展.最后提出该领域未来的研究重点.参147
|
| [29] |
Nitrogen additions and microbial biomass: a meta—analysis of ecosystem studies [J].https://doi.org/10.1111/j.1461-0248.2008.01230.x URL PMID: 18673384 [本文引用: 1] 摘要
Nitrogen (N) enrichment is an element of global change that could influence the growth and abundance of many organisms. In this meta-analysis, I synthesized responses of microbial biomass to N additions in 82 published field studies. I hypothesized that the biomass of fungi, bacteria or the microbial community as a whole would be altered under N additions. I also predicted that changes in biomass would parallel changes in soil CO2 emissions. Microbial biomass declined 15% on average under N fertilization, but fungi and bacteria were not significantly altered in studies that examined each group separately. Moreover, declines in abundance of microbes and fungi were more evident in studies of longer durations and with higher total amounts of N added. In addition, responses of microbial biomass to N fertilization were significantly correlated with responses of soil CO2 emissions. There were no significant effects of biomes, fertilizer types, ambient N deposition rates or methods of measuring biomass. Altogether, these results suggest that N enrichment could reduce microbial biomass in many ecosystems, with corresponding declines in soil CO2 emissions.
|

/
| 〈 |
|
〉 |
{kind=link}
{kind=link}