王晓东 , 刘惠清
, 刘惠清
东北师范大学城市与环境科学学院, 吉林 长春 130024
WANG Xiao-dong, LIU Hui-qing
中图分类号: K903
文献标识码: A
文章编号: 1000-0690(2012)02-0199-08
通讯作者:
收稿日期: 2011-04-6
修回日期: 2011-07-10
网络出版日期: 2012-02-20
版权声明: 2012 《地理科学》编辑部 本文是开放获取期刊文献,在以下情况下可以自由使用:学术研究、学术交流、科研教学等,但不允许用于商业目的.
基金资助:
作者简介:
作者简介:王晓东(1971-),男,辽宁盘锦人,博士研究生,主要从事自然地理研究。E-mail :wangxd219@nenu.edu.cn
展开
摘要
选取长白山北坡岳桦(Betula ermanii)苔原交错带3个不同坡向(东北坡、东南坡、东坡)的岳桦径级结构为研究对象,分析坡向在岳桦种群动态与气候变化之间关系的异同。结果表明:三坡向的岳桦种群扩张和气温高低存在明显差异。14个生态气候指标与对应年份内繁殖且存活下来的岳桦棵数的逐步回归分析和相关分析表明三坡向岳桦种群扩张是对气温变化的积极响应,但响应有差异:东坡>东南坡>东北坡。不同坡向岳桦种群扩张的差异是对气温变化差异的响应。
关键词:
Abstract
To discover the relation between Betula ermanii population dynamics and climate change on treeline ecotone of Changbai Mountains, 53 plots were selected on three different slope aspects (northeast, southeast, east slopes) through treeline ecotone on north slope of Changbai Mountains. Distributing character of the diameter-class (diameter at breast height (DBH)) structure of shrub and single-trunk forms were described by four aggregation indices. Trend analysis of distributing character was carried through by unitary linearity equations. Different reflection of the dynamics response of Betula ermanii population to climate change on different slope aspects was analyzed by stepwise regression and correlation analysis. The results are as follows:the results of trend analysis of the diameter-class distributing (slopes of equations are negative) shows the capacity expansion rate of shrub form is east slopes>southeast slopes>northeast slopes and that of single trunk form is northeast slopes>east slopes>southeast slopes. The population expansion is leapfrog and pulsatile from low to high (p>0.01).The result of One-way ANOVA displays that daily mean temperature on three slopes are significant differences (p<0.05) and the temperature change has synchronism. The results of correlation analysis between annual stem recruitment of Betula ermanii population and 14 ecological preferences of air temperature (1953 - 2008) show that the different response of population dynamics to climate change caused by different temperature on three slopes. The difference of climate change lead to different expansion of Betula ermanii population on different positions.
Keywords:
从山地郁闭林到树种线间的过渡带称为高山林线[1,2],对全球气候变化高度敏感[3,4],探讨林线变动和气候变化的关系已经成为现在研究的热点[5,6]。许多学者在分析林线变动对气候变化的响应方式时注意到地形因子对这种关系有着一定影响,因此把地形要素纳入到研究体系中[7]。如分析制约林线植物生长的气温、土壤等生态因子时揭示地形的特殊影响[8],应用生态模型认识林线气候—植被关系涉及各种地形要素[9],使用GIS技术分析林线动态时显示地形要素是重要的生态因子[10],对影响太白红杉林分布的控制因子进行分析时发现坡向对林线变动有显著影响[11]。在风雪对林线上缘景观变化影响的研究中重点强调地形因子的作用[12]。然而,地形因子对林线变动与气候变化关系影响的研究成果很少[4],特别是坡向差异对这种响应关系的制约还需深入分析。
本文以坡向分异为切入点,以数学模型、单因素方差、相关分析等方法对气温测量拟合,与野外样方调查和树轮技术方法所得数据综合分析,对长白山北坡林线不同坡向岳桦(Betula ermanii)种群径级结构进行统计分析,揭示种群动态,判断种群扩张与气候变化的关系。进一步分析由于地形坡向分异造成的岳桦种群差异性扩张及其影响因子。
长白山自然保护区位于中国东北地区吉林省东南部,深受太平洋影响,与朝鲜民主主义人民共和国毗邻。地理位置是41°41′49″N~42°25′18″N,127°42′55″E ~128°16′48″E。属于具有季风影响的温带大陆性山地气候,可划分中温带、寒温带和高山亚寒带3个气候带[13]。随着地势的增高,自下而上构成了以地带性针阔混交林为基带的几个垂直植被带[14]。红松(Pinus koraiensis)针阔混交林,红松占据上层林冠,分布在海拔700~1 100 m范围内;针叶林分布在海拔1 100~1 700 m范围内,是以鱼鳞云杉(Picea jezoensis var. komarovii)和臭冷杉(Abies nephrolepis)占优势的群落;在海拔1 700~2 000 m范围内是以岳桦为主的纯林;2 000 m以上为高山苔原带,生长具有旱生形态的湿冷植物[15,16]。
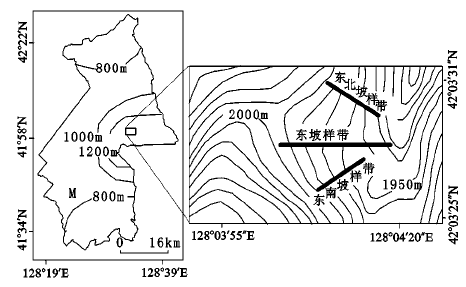
长白山北坡林线岳桦的海拔为1 900~2 000 m 左右,局部地区可以上升至2 100 m以上[17]。林线生境风大寒冷,不适于其他树种生长,遂使岳桦成为优势种[18]。研究区位于长白山北坡岳桦与苔原交错带,地理位置为42°3′25″N ~42°3′31″N,128°3′55″E ~128°4′20″E,是长白山北坡林线的一部分,海拔为1 950~2 070 m,林线分别刚好在地形的不同部位通过。
2008年9月采用样方调查法在长白山北坡一段林线经过的区域内(研究区)选择地域相连的3个坡,即东坡,东北坡和东南坡的不同坡向上沿林线边缘自高而低连续取样一直到谷底的郁闭林中(郁闭度>0.2)(图1),在东南坡采集15个样方,在东北坡采集13个样方,在东坡采集25个样方。用GPS测定样地的经纬度和海拔,用坡度仪测定坡度,样方的大小为10 m×10 m,主要调查样方内灌丛状和乔木状岳桦的个体数量和记录胸径的大小。胸径用卷尺测量(精确到0.1 cm),树高小于2 m用测绳,大于2 m用平板仪测算;树高大于1.5 m精确到0.1 m,小于1.5 m精确到0.01 m。
图 1 长白山北坡林线样地
Fig. 1 Sampling plots within the alpine treeline on the north-facing slope of Changbai Mountains
在每一样方内选择典型岳桦个体(涵盖到每一径级)做树轮分析,测定方法采用树芯和树盘两种方式:径级大于3 cm,取树芯,共得89个;在98棵受到破坏与死亡的小树上共取127个树盘。这些小树包括一些风折和冻死的树,人为及动物踩踏破坏的树,以及十几棵采样时不慎踩折的树。在距地表0.3 m的树干处取2个树芯;并在距地表0.3 m的树干处取树盘;径级小于1 cm、树高小于1 m的则直接在基部取样。取得的树芯和树盘经干燥打磨后在仪器(LA-S型植物年轮分析系统:2亿像素)上交叉定年[19],获取年轮数据。按照树轮分析法所提取出的样地内岳桦各个径级所对应的生长年龄,按岳桦的生长规律[20],曲仲湘的分级原理[21]与龄级划分的方法[22],分别在3个样地内建立径级—年龄(树高≥1.5 m)和年龄—树高(树高<1.5 m)的数学模型,最后分别将3个样地内岳桦径级结构转化为对应的年龄结构(表 1),参照样方调查数据推算特定年份内繁殖并且存活下来的岳桦棵数。
表 1 样地内岳桦年龄和径级、树高数学模型
Table 1 The age-class and age-height regressions of Betula ermanii in quadrats
 样地 样地 | 树高大于1.5 m | 树高小于1.5 m | ||
|---|---|---|---|---|
| 灌丛状 | 乔木状 | 灌丛状 | 乔木状 | |
| 东坡 | y=e(2.5745+0.0926x) | y=e(2.4836+0.1541x) | y=7.0125t0.8236 | y=6.6588t0.7916 |
| 东北坡 | y=e(2.6442+0.1132x) | y=e(2.5467+0.1081x) | y=7.2705t0.9272 | y=6.5295t0.9011 |
| 东南坡 | y=e(2.5735+0.0819x) | y=e(2.5435+0.1198x) | y=6.6524t0.9358 | y=6.8814t0.7921 |
注:y为树龄,x为胸径,t为树高。
在3个样地内把6个自动测温仪(HOBO-Temp data-logger)分别放置在海拔相同的位置上(1 980 m)测量3个坡上的气温(距地面1.2 m)与地表温度(距地面0 m),记录时间间隔为1 h,测温时间从2008年5月1日到2009年5月1日,统计实地监测的气温和地温数据,逐日、逐月计算该时段内3个坡上的日平均气温与月平均气温。
利用2008年天池气象站(2 624 m)的气温数据和长白山北坡垂直递减率[23]推算出1 980 m处气温值(日平均气温,月平均气温),推算结果与实地监测得到的日平均气温与月平均气温数据进行相关分析(p <0.05),显示3个坡上气温变化具有高度一致性,因此可用天池气象站气温数据经过校正来代替这些坡上气温数据。于是用天池台站多年气象资料(1953~2008年)拟推得到3个坡上56 a的14个生态气候指标,即1月、5月、7月、9月平均气温、年平均气温,∑t≥0℃、∑t≥10℃,5月、7月、9月、6~8月、5~9月降水量、温暖指数(WI)和湿润指数(HI)[24]。
根据径级结构体现年龄结构原理[25],采用4个聚集度指数(丛生度I[26]、聚块度M*/m[27]、Cassie指标CA[28]、扩散指标C[29])分析岳桦种群的径级结构来认识种群的动态特征。由于测定分布格局的指标很多,各指标计算结果有不一致地方,所以采用多指标对测定结果的综合分析。另外,这些分布型指数能方便地测度径级分布的聚集性[30],是揭示种群时间系列变化的有力工具[31]。指标值越大,不同径级间个体数量差异越明显[32],种群扩张态势越显著。
单因素方差分析模块下的多重比较,尤其是LSD方法经常用于检验统计量的观测值和概率p值来判断样本间差异比较和显著性程度。所以,应用这种统计分析方法分析检验在地域接近的3个不同坡向上气温高低(日平均气温)和差异的显著性程度。
先采用线型方程对聚集度指标进行趋势拟合,然后将3个坡上校订拟推出的14个生态气候指标(1953~2008年)和对应年份内繁殖并且存活下来的岳桦棵数进行逐步回归分析判断岳桦种群动态和气候变化的关系,最后用相关分析揭示由于地形分异形成的岳桦种群动态对气候变化响应的差异性。
径级结构是年龄结构的反映[33],用岳桦种群径级结构可以分析种群变化。林线处岳桦种群的生活型有灌丛状和乔木状两种,灌丛状岳桦通过无性繁殖直接从母体内得到养分、乔木状岳桦是通过种子繁殖靠自身能量生长[34]。不同的生长方式对气候变化的响应不同[35],因此将岳桦分为灌丛状和乔木状分别阐述。
由表2可知,岳桦种群径级结构在3个坡上都是聚集分布(径级分布p<0.01, I>0, M*/m>0,
表2 岳桦种群径级分布的聚集度指标
Table 2 The aggregation indices of Betula ermanii diameter configuration
| 样地编号 | 样方编号 | 灌丛状 | 乔木状 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| TD1 | I | M*/m | CA | C | TD2 | I | M*/m | CA | C | ||
| 东 坡 | 1 | 25 | 20.173 | 21.980 | 20.980 | 21.173 | 12 | 11.000 | 18.417 | 17.417 | 12.000 |
| 2 | 22 | 1.996 | 3.359 | 2.359 | 2.996 | 10 | 9.000 | 18.100 | 17.100 | 10.000 | |
| 3 | 86 | 25.381 | 8.673 | 7.673 | 26.381 | 53 | 23.759 | 9.517 | 8.517 | 24.759 | |
| 4 | 34 | 3.513 | 3.686 | 2.686 | 4.513 | 61 | 25.288 | 8.877 | 7.877 | 26.288 | |
| 5 | 19 | 2.101 | 3.764 | 2.764 | 3.101 | 32 | 12.066 | 8.164 | 7.164 | 13.066 | |
| 6 | 11 | 1.113 | 3.630 | 2.630 | 2.113 | 74 | 35.884 | 10.214 | 9.214 | 36.884 | |
| 7 | 46 | 9.911 | 6.602 | 5.602 | 10.911 | 58 | 33.669 | 12.029 | 11.029 | 34.669 | |
| 8 | 67 | 27.660 | 11.734 | 10.734 | 28.660 | 58 | 25.370 | 9.311 | 8.311 | 26.370 | |
| 9 | 90 | 28.195 | 9.145 | 8.145 | 29.195 | 60 | 17.411 | 6.514 | 5.514 | 18.411 | |
| 10 | 34 | 16.421 | 13.557 | 12.557 | 17.421 | 48 | 19.336 | 8.654 | 7.654 | 20.336 | |
| 11 | 87 | 12.172 | 4.638 | 3.638 | 13.172 | 80 | 58.971 | 15.006 | 14.006 | 59.971 | |
| 12 | 75 | 13.486 | 5.675 | 4.675 | 14.486 | 49 | 33.567 | 14.016 | 13.016 | 34.567 | |
| 13 | 37 | 5.081 | 4.571 | 3.571 | 6.081 | 76 | 27.889 | 7.972 | 6.972 | 28.889 | |
| 14 | 49 | 7.801 | 5.139 | 4.139 | 8.801 | 118 | 24.773 | 4.989 | 3.989 | 25.773 | |
| 15 | 62 | 14.670 | 7.152 | 6.152 | 15.670 | 77 | 16.724 | 5.127 | 4.127 | 17.724 | |
| 16 | 61 | 5.511 | 3.349 | 2.349 | 6.511 | 2 | -0.056 | 0.472 | -0.528 | 0.944 | |
| 17 | 0 | 0.000 | 1.000 | 0.000 | 1.000 | 44 | 22.609 | 10.763 | 9.763 | 23.609 | |
| 18 | 18 | 3.942 | 6.694 | 5.694 | 4.942 | 11 | 2.707 | 5.676 | 4.676 | 3.707 | |
| 19 | 83 | 16.342 | 6.119 | 5.119 | 17.342 | 12 | 3.259 | 6.161 | 5.161 | 4.259 | |
| 20 | 217 | 43.274 | 6.185 | 5.185 | 44.274 | 22 | 4.975 | 5.296 | 4.296 | 5.975 | |
| 21 | 30 | 5.912 | 6.124 | 5.124 | 6.912 | 8 | 3.569 | 9.477 | 8.477 | 4.569 | |
| 22 | 9 | 2.684 | 8.755 | 7.755 | 3.684 | 6 | 3.241 | 11.262 | 10.262 | 4.241 | |
| 23 | 0 | 0.000 | 1.000 | 0.000 | 1.000 | 6 | 2.185 | 7.920 | 6.920 | 3.185 | |
| 24 | 9 | 6.151 | 18.770 | 17.770 | 7.151 | 11 | 5.010 | 9.654 | 8.654 | 6.010 | |
| 25 | 39 | 5.893 | 4.929 | 3.929 | 6.893 | 73 | 19.540 | 6.086 | 5.086 | 20.540 | |
| 东 北 坡 | 1 | 0 | 0.000 | 1.000 | 0.000 | 1.000 | 4 | 3.000 | 15.250 | 14.250 | 4.000 |
| 2 | 63 | 20.334 | 9.392 | 8.392 | 21.334 | 0 | 0.000 | 1.000 | 0.000 | 1.000 | |
| 3 | 0 | 0.000 | 1.000 | 0.000 | 1.000 | 64 | 33.214 | 10.860 | 9.860 | 34.214 | |
| 4 | 18 | 9.951 | 15.374 | 14.374 | 10.951 | 32 | 17.344 | 11.298 | 10.298 | 18.344 | |
| 5 | 33 | 7.418 | 6.845 | 5.845 | 8.418 | 45 | 44.000 | 19.578 | 18.578 | 45.000 | |
| 6 | 5 | 1.504 | 8.821 | 7.821 | 2.504 | 29 | 8.345 | 6.467 | 5.467 | 9.345 | |
| 7 | 0 | 0.000 | 1.000 | 0.000 | 1.000 | 23 | 16.493 | 14.625 | 13.625 | 17.493 | |
| 8 | 0 | 0.000 | 1.000 | 0.000 | 1.000 | 78 | 35.860 | 9.735 | 8.735 | 36.860 | |
| 9 | 11 | 1.680 | 4.971 | 3.971 | 2.680 | 51 | 11.876 | 5.424 | 4.424 | 12.876 | |
| 10 | 12 | 2.853 | 7.182 | 6.182 | 3.853 | 58 | 21.366 | 7.999 | 6.999 | 22.366 | |
| 11 | 0 | 0.000 | 1.000 | 0.000 | 1.000 | 53 | 22.126 | 8.932 | 7.932 | 23.126 | |
| 12 | 11 | 0.924 | 3.183 | 2.183 | 1.924 | 79 | 43.581 | 11.482 | 10.482 | 44.581 | |
| 13 | 25 | 4.032 | 5.193 | 4.193 | 5.032 | 38 | 37.000 | 19.500 | 18.500 | 38.000 | |
| 东 南 坡 | 1 | 88 | 9.851 | 3.911 | 2.911 | 10.851 | 21 | 3.614 | 4.270 | 3.270 | 4.614 |
| 2 | 45 | 7.854 | 5.538 | 4.538 | 8.854 | 61 | 16.878 | 6.257 | 5.257 | 17.878 | |
| 3 | 44 | 2.487 | 2.470 | 1.470 | 3.487 | 20 | 3.483 | 4.309 | 3.309 | 4.483 | |
| 4 | 82 | 16.596 | 6.262 | 5.262 | 17.596 | 10 | 0.978 | 2.858 | 1.858 | 1.978 | |
| 5 | 21 | 3.063 | 4.792 | 3.792 | 4.063 | 33 | 5.067 | 3.918 | 2.918 | 6.067 | |
| 6 | 97 | 8.147 | 3.184 | 2.184 | 9.147 | 55 | 11.125 | 4.843 | 3.843 | 12.125 | |
| 7 | 54 | 8.164 | 4.931 | 3.931 | 9.164 | 1 | 0.000 | 1.000 | 0.000 | 1.000 | |
| 8 | 104 | 5.620 | 2.405 | 1.405 | 6.620 | 63 | 11.668 | 4.519 | 3.519 | 12.668 | |
| 9 | 86 | 10.749 | 4.250 | 3.250 | 11.749 | 1 | 0.000 | 1.000 | 0.000 | 1.000 | |
| 10 | 18 | 1.631 | 3.356 | 2.356 | 2.631 | 31 | 25.982 | 16.925 | 15.925 | 26.982 | |
| 11 | 69 | 8.554 | 4.223 | 3.223 | 9.554 | 4 | 3.000 | 15.250 | 14.250 | 4.000 | |
| 12 | 41 | 1.850 | 2.173 | 1.173 | 2.850 | 4 | 1.417 | 7.729 | 6.729 | 2.417 | |
| 13 | 44 | 2.298 | 2.358 | 1.358 | 3.298 | 10 | 0.344 | 1.654 | 0.654 | 1.344 | |
| 14 | 70 | 2.262 | 1.840 | 0.840 | 3.262 | 34 | 1.147 | 1.641 | 0.641 | 2.147 | |
| 15 | 0 | 0.000 | 1.000 | 0.000 | 1.000 | 35 | 4.083 | 3.216 | 2.216 | 5.083 | |
1) 灌丛状方式径级分布的对比分析。一元线型方程趋势分析(表 3)表明,岳桦种群以灌丛状方式沿海拔从低向高不断扩张(斜率都是负值)。扩张能力和速率大小是东坡>东南坡>东北坡(东坡、东南坡、东北坡4个聚集度指标的均值是9.112>4.727>4.401;对应斜率的均值是-0.16<-0.39<-0.41),岳桦以灌丛方式的上侵不是平移匀速推进的,而是以蛙跳脉动式向上拓展(p>0.01),脉动幅度是东坡>东北坡>东南坡(东坡、东北坡、东南坡的p分别是0.501>0.346>0.021;R2分别是0.021<0.367>0.097)。
2) 乔木状方式径级分布的对比分析。一元线型方程的趋势分析(表 3)表明岳桦种群以乔木状方式的扩张态势较复杂(斜率正负值都有)。扩张能力的大小是东北坡>东坡>东南坡(东北坡、东坡、东南坡4个聚集度指标的均值是16.782 8>13.428 4>5.605 9),扩张态势是东北坡在下部扩张强烈,东坡,东南坡在上部扩张明显且东坡>东南坡(东北坡、东坡、东南坡聚集度方程对应的斜率均值是1.065、-0.550、-0.100)。岳桦以乔木方式的上侵也不是平移匀速推进的,而是以蛙跳脉动式向上拓展(p>0.01),脉动幅度是东南坡>东北坡>东坡(东南坡,东北坡,东坡的p均值分别是0.631>0.362>0.024;R2均值分别是0.021<0.138<0.208)。
表3 岳桦种群聚集度线性趋势分析
Table 3 The linear trend analysis of Betula ermanii diameter configuration
| 生长方式 | 样地编号 | 方程与参数 | I | M*/m | CA | C |
|---|---|---|---|---|---|---|
| 灌丛状 | 东坡 | 方程 | y=-0.22x+14.05 | y=-0.09x+8.21 | y=-0.09x+7.21 | y=-0.22x+15.05 |
| p | 0.473 | 0.528 | 0.528 | 0.473 | ||
| R2 | 0.023 | 0.018 | 0.018 | 0.023 | ||
| 东北坡 | 方程 | y=-0.59x+7.87 | y=-0.23x+6.70 | y=-0.23x+5.69 | y=-0.59x+8.87 | |
| p | 0.188 | 0.504 | 0.504 | 0.188 | ||
| R2 | 0.152 | 0.042 | 0.042 | 0.152 | ||
| 东南坡 | 方程 | y=-0.56x+10.41 | y=-0.22x+5.27 | y=-0.22x+4.27 | y=-0.56x+11.41 | |
| p | 0.034 | 0.008 | 0.008 | 0.034 | ||
| R2 | 0.301 | 0.433 | 0.433 | 0.301 | ||
| 乔木状 | 东坡 | 方程 | y=-0.82x+28.33 | y=-0.27x+12.68 | y=-0.27x+11.68 | y=-0.82x+29.33 |
| p | 0.032 | 0.015 | 0.015 | 0.032 | ||
| R2 | 0.184 | 0.231 | 0.231 | 0.184 | ||
| 东北坡 | 方程 | y=1.94x+9.06 | y=0.19x+9.58 | y=0.19x+8.58 | y=1.94x+10.06 | |
| p | 0.078 | 0.646 | 0.646 | 0.078 | ||
| R2 | 0.256 | 0.02 | 0.02 | 0.256 | ||
| 东南坡 | 方程 | y=-0.29x+8.25 | y=0.10x+4.46 | y=0.10x+3.46 | y=-0.29x+9.25 | |
| p | 0.533 | 0.728 | 0.728 | 0.533 | ||
| R2 | 0.031 | 0.01 | 0.01 | 0.031 |
为分析在不同坡向上岳桦种群差异性扩张的影响因素,对3个坡上气温特点和差异进行分析。多重比较分析的结果(表4)显示:虽然地域相连,海拔相近(1 950~2 000 m),但不同坡上温度高低差异显著(除地温的东坡与东南坡的差异外,其他指标p<0.05),气温和地温高低排序是东北坡>东坡>东南坡。
表4 不同坡上的气温和地温的差异
Table 4 The differences among different slopes on air temperatures and ground temperatures
| 坡向差异 | 均值差(℃) | p | |
|---|---|---|---|
| 气温 | 东坡 东南坡 | 1.88* | 0.003 |
| 东北坡 东南坡 东坡 | 3.52* | 0.000 | |
| 1.65* | 0.009 | ||
| 地温 | 东坡 东南坡 | 1.01 | 0.194 |
| 东北坡 东南坡 东坡 | 3.80* | 0.000 | |
| 2.79* | 0.000 |
长白山地区气候经历明显变化[36],变暖趋势明显[37~40]。按照林线树木生长不但与温度有关,而且与降水紧密相连的结论[41],采用相关和逐步回归分析揭示林线岳桦种群变动与气候变化的关系,同时进一步认识岳桦种群变动对气候变化响应方式在不同坡向上的差异性表现。
1) 种群变动对气候变化的响应。根据逐步回归分析结果(表 5)显示,纳入到回归方程中的气温指标个数大于降水指标(气温因子35个>降水因子12个),纳入指标的显著性水平气温指标个数也高于降水(p<0.05:气温因子18个>降水因子2个),表明气温变化是种群变动的控制因子,由于降水充足(1 100 mm),降水变化不是林线进退的制约因子,然而,个别月份因降水量少(如9月降水量占全年7.7%),对种群扩张有一定影响(9月降水量纳入所有方程中,其中达到显著性个数2个)。
表5 生态气候指标与对应年份内繁殖并且存活下来的岳桦棵数逐步回归分析
Table 5 Stepwise regression between annual stem recruitment and ecological preferences
| 东坡 | 灌丛状 | 东南坡 | 东坡 | 乔木状 | 东南坡 | ||
|---|---|---|---|---|---|---|---|
| 东北坡 | 东北坡 | ||||||
| 自变量 | x1 | -0.10 | |||||
| x2 | -0.26 | -0.25 | -0.23 | -0.29 | |||
| x3 | |||||||
| x4 | 0.35* | 0.16 | 0.37* | 0.26 | 0.17 | 0.30 | |
| x5 | -0.06 | ||||||
| x6 | 0.22 | ||||||
| x7 | 0.11 | ||||||
| x8 | 0.26 | 0.22 | 0.34* | 0.29 | 0.33* | ||
| x9 | 0.21 | 0.08 | 0.19 | 0.24 | 0.23 | 0.36* | |
| x10 | 0.26 | 0.25 | 0.26 | 0.17 | 0.34* | ||
| x11 | 0.53** | 0.09 | 0.39 | 0.51** | 0.44** | 0.54** | |
| x12 | 0.56** | 0.57** | 0.59** | 0.45** | 0.34* | 0.46** | |
| x13 | 0.21 | 0.09 | 0.38* | 0.14 | 0.31* | ||
| x14 | -0.31 | -0.14 | -0.44** | -0.25 | -0.12 | -0.37* | |
| 模型 检验 | y1 | 0.0001 | 0.0120 | 0.0001 | 0.0002 | 0.0023 | 0.0001 |
| y2 | 1.3875 | 1.5347 | 1.0522 | 1.4012 | 1.2861 | 1.3336 | |
相关分析结果(表6)显示在不同坡上繁殖并且存活下来的岳桦棵数(灌丛状与乔木状)和气温指标的相关性高于降水指标[显著性指标(p<0.05)个数:气温24个>降水5个],进一步验证岳桦种群扩张是对气候变暖的积极响应[42]。对9月降水量指标相关性较高[显著性指标(p<0.05)个数为5个]则有利证明由于降水季节分配不均(p<0.05),9月降水量不足是种群扩张的影响因子。
表6 生态气候指标与对应年份内繁殖并且存活下来的岳桦棵数的相关分析
Table 6 Correlation analysis between annual stem recruitment and ecological preferences
| 相关 系数 | 东坡 | 灌丛状 | 东南坡 | 东坡 | 乔木状 | 东南坡 |
|---|---|---|---|---|---|---|
| 东北坡 | 东北坡 | |||||
| x1 | 0.01 | -0.02 | -0.1 | 0.01 | 0.01 | -0.02 |
| x2 | 0 | -0.2 | -0.02 | -0.02 | -0.07 | -0.01 |
| x3 | 0.48** | 0.08 | 0.30* | 0.46** | 0.39** | 0.45** |
| x4 | 0.23 | -0.06 | 0.12 | 0.2 | 0.14 | 0.2 |
| x5 | 0.06 | -0.12 | 0.03 | 0.03 | -0.01 | 0.04 |
| x6 | 0.09 | -0.04 | 0.03 | 0.08 | 0.08 | 0.08 |
| x7 | 0.13 | 0.18 | 0.15 | 0.11 | 0.11 | 0.1 |
| x8 | 0.17 | 0.18 | 0.24 | 0.14 | 0.14 | 0.1 |
| x9 | 0.39** | 0.21 | 0.36* | 0.41** | 0.42** | 0.32* |
| x10 | 0.41** | 0.30* | 0.42** | 0.38** | 0.39** | 0.35* |
| x11 | 0.51** | 0.12 | 0.44** | 0.40** | 0.54** | 0.42** |
| x12 | 0.48** | 0.51** | 0.54** | 0.40** | 0.48** | 0.39** |
| x13 | 0.27 | 0.19 | 0.11 | 0.30* | 0.32* | 0.13 |
| x14 | -0.07 | -0.18 | -0.09 | -0.1 | -0.13 | -0.04 |
2) 坡向分异造成的差异性响应。对岳桦种群动态分析时发现径级分布呈聚集状态且3个坡上还存在一定差异,反映岳桦种群变动深受气候变化的影响而且在不同地形位置上种群动态对气候变化的响应程度是不同的。
灌丛状种群变动对气候变化响应的相关分析结果显示三坡相关系数的大小(相关系数均值:东坡0.30>东南坡0.29>东北坡0.21)与三坡灌丛方式扩张能力的大小相同(表6),表明三坡灌丛状种群变动差异与温度差异有关,东坡温度比东南坡高促使种群扩张更强,而东北坡扩张能力最弱是由于温度持续升高促使在种群扩张的过程中生长方式转变,灌丛方式反而不积极。
乔木状种群变动对气候变化响应的相关分析结果显示乔木方式对气候条件要求更严格(显著性相关个数:乔木状14个>灌丛状10个)造成林线迅速升温使乔木方式的活力增强,相关系数的大小(相关系数均值:东北坡0.31>东坡0.28>东南坡0.24)与三坡乔木方式扩张能力的大小相同(表6),在同样升温条件下,温度越高的位置岳桦越以扩散能力强的种子繁殖方式来尽快实现种群扩张。
1) 岳桦种群的聚集度分析显示岳桦种群(灌丛和乔木生长方式)径级呈聚集分布,表明林线岳桦种群在不断扩张变化。
2) 对样地内计算得到的聚集度进行线性方程的趋势分析表明岳桦种群沿海拔梯度从低向高上侵(斜率为负),但这种上侵不是平移推进的,而是以蛙跳脉动形式来实现种群的扩张(p>0.01)。
3) 通过对线性方程及有关参数分析表明在不同坡向上岳桦种群(乔木与灌丛生长方式)的扩张是有差异的。
4) 对实测日均温数据在3个坡上的差异性单因素方差分析结果表明不同坡向的气温高低存在显著差异(p<0.05)。
5) 在不同坡上生态气候指标与繁殖并且存活下来的岳桦棵数的相关和逐步回归分析结果显示岳桦种群动态与气候变化密切关系,岳桦种群的上侵是对气候变化的响应,而且这种响应因坡向分异而出现明显区别。
在研究时发现同一坡内差异也比较大,可能与海拔高度,坡度,微地形等差异和气候年际变化有关,由于本文是以地形差异中的坡向分异为切入点对种群扩张的原因进行探讨,对于同坡内的差异在以后研究中逐步深入分析,在此不再论述。
由于缺乏长序列的气象资料,没有在整个岳桦生长期内建立年龄-气候间的对应关系,可能对分析结果带来一些偏差(相关系数偏低)。林线变动是对各种影响因素的综合响应,而本文只对气温降水进行分析,没有考虑风,积雪,光照等其它气候因子。对地形中的坡度,坡长,曲率,地面组成物质,土壤的理化性质,养分的差异也没有纳入进来,可能使分析结果造成一些影响。
The authors have declared that no competing interests exist.
| [1] |
Alpine Plant Life:Functional Plant Ecology of High Mountain Ecosystems [M]. |
| [2] |
A world 2 wide study of high altitude treeline temperatures [J]. |
| [3] |
Variability in an edaphic indicator in alpine tundra [J]. |
| [4] |
Spatial pattern of subalpine forest-alpine grassland ecotones in the Spanish Central Pyrenees [J]. |
| [5] |
1998~2008年中国东部沿海植被覆盖变化特征 [J]. |
| [6] |
Growing-season microclimate variability within an old-growth Douglas-fir forest [J]. |
| [7] |
Implications of patterns of carbon pools and fluxes across a semiarid environmental gradient [J]. |
| [8] |
Kinetics of plant growth and metabolism [J]. |
| [9] |
DawsonPotential effects of climate change on plant communities in three montane nature reserves in Scotland,UK [J]. |
| [10] |
Mapping,modeling,and visualization of the influences of geomorphic processes on the alpine treeline ecotone,Glacier National Park,MT,USA [J]. |
| [11] |
秦岭太白红杉林分布及太白山高山林线特征的定量分析 [J]. |
| [12] |
Observing, modelling, and validating snow redistribution by wind in a Wyoming upper treeline landscape [J]. |
| [13] |
2000~2008年长白山地区植被覆盖变化对气候的响应研究 [J]. |
| [14] |
长白山岳桦林化学元素生物地球化学分析 [J]. |
| [15] |
长白山北侧的自然景观带 [J]. |
| [16] |
长白山北坡苔原/岳桦景观边界的定量检测 [J]. |
| [17] |
长白山岳桦种群格局的地统计学分析 [J]. |
| [18] |
长白山林线交错带形状与木本植物向苔原侵展和林线动态的关系 [J]. |
| [19] |
Comper-assisted quality control in tree-ring dating and measurement [J]. |
| [20] |
长白山上岳桦林的调查研究 [C]// |
| [21] |
长白山岳桦苔原过渡带动态与气候变化 [J]. |
| [22] |
长白山北坡岳桦种群结构及动态分析 [J]. |
| [23] |
|
| [24] |
东北农牧交错区水分条件及其对植被分布的影响 [J]. |
| [25] |
Relationships of climate change and tree ring of Betula ermanii tree line forest in Changbai Mountain [J]. |
| [26] |
元宝山南方红豆杉克隆种群分布格局的分形特征 [J]. |
| [27] |
长白山林区核桃楸种群分布格局研究 [J]. |
| [28] |
濒危植物元宝山冷杉种群结构与分布格局 [J]. |
| [29] |
子午岭油松林灌木层主要树种的空间分布格局和种间关联性研究 [J]. |
| [30] |
陕西府谷自然保护区杜松种群的空间分布格局及其动态植物 [J]. |
| [31] |
珍稀濒危植物桫椤种群结构与动态研究应用 [J]. |
| [32] |
绿盲蝽越冬卵在枣树上的空间分布型研究 [J]. |
| [33] |
Climatic control of primary forest structure and DBH-height allometry in Northeast China [J]. |
| [34] |
养分供应对3种舞花姜属植物繁殖方式的影响 [J]. |
| [35] |
长白山北坡林线岳桦种群与土壤关系的研究 [J]. |
| [36] |
1951~2005年中国大陆冬季温度变化过程的区域差异 [J]. |
| [37] |
基于FORCCHN的未来东北森林生态系统碳储量模拟 [J]. |
| [38] |
长白山阔叶红松林近22年的气候动态 [J].
|
| [39] |
大气增温对长白山林木直径生长的影响 [J].
|
| [40] |
Dendroclimatic analysis of Betula ermanii forests at their upper limit of distribution in Changbai Mountain, Northeast China [J]. |
| [41] |
高山树线交错带的景观格局与生态过程 [J]. |
| [42] |
长白山北坡岳桦林线变动的水热条件分析 [J]. |

/
| 〈 |
|
〉 |
{kind=link}
{kind=link}